




Фокин С.Ю. Звуковая коммуникация и поведение обыкновенного фазана при искусственном содержании // Искусственное разведение фазанов. Сборник научных трудов ЦНИЛ Главохоты РСФСР. Москва, 1983. С. 158-168.
Специальные исследования последних лет в области прикладной биоакустики показали принципиальную возможность управления поведением птиц при искусственном содержании. Трудность же заключается в недостаточном знании поведения и основных звуковых реакций многих видов птиц - объектов дичеразведения. К числу таких видов относится и обыкновенный фазан.
Целью нашей работы было изучение птенцового, брачного и родительского поведения обыкновенного фазана, его акустической сигнализации и возможность ее использования в фазановодстве. Эксперименты проводились в дичепитомнике ЦНИЛ Главохоты РСФСР. Для записи голосов птиц использовали магнитофон "Электроника-321" с микрофоном МД-200. Спектрально-временной анализ звуковых сигналов проводили на динамическом спектроанализаторе "Спектр-1".
Звуковая сигнализация в брачный период
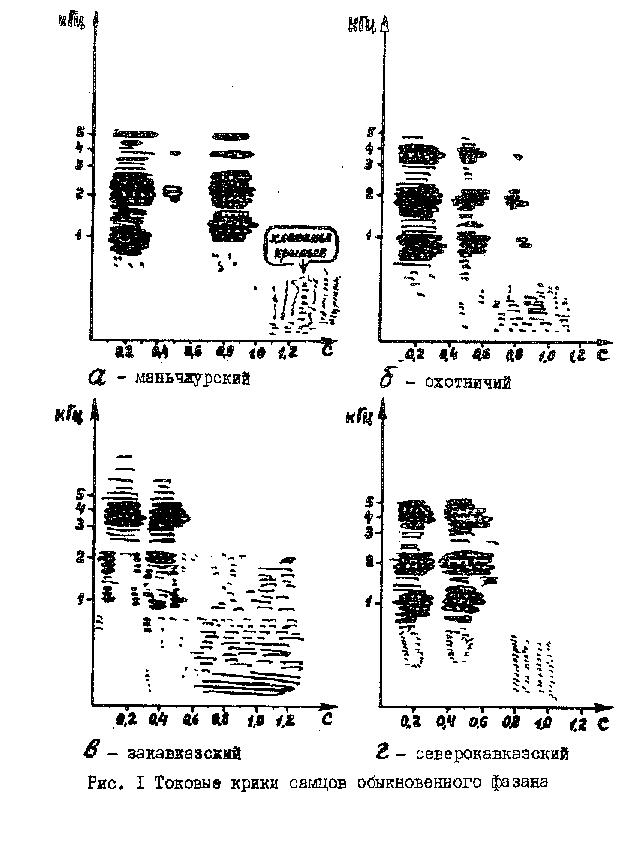
В брачный период можно выделить 4основных типа звуковыхсигналов. Токовый крик. Во время брачных демонстраций самец принимает характерную "выпрямленную" позу, вытягивая шею и подаваясь всем корпусом вверх. После этого следует быстрый взмах крыльями, резкий двусложный крик, по окончании которого самец громко хлопает крыльями. Токовый крик вызывает аналогичный ответ у соседних самцов (по типу петушиной переклички), а также активирует брачное поведение самок, которые в ответ на сигнал петуха издают тихие комфортные сигналы. Однако токовый крик не всегда предшествует спариванию. Нами отмечено, что "песенная" активность самца не коррелирует с числом "садок". Часто бывает, что наиболее молчаливые петухи спариваются чаще, чем голосистые. В то же время самки, сидящие в одной вольере с акустически активным петухом, проявляют большее к нему предпочтение, чем фазанки, находящиеся с "молчуном". Этот эффект, очевидно, можно использовать для акустической стимуляции яйценоскости фазанок. Интересно, что токовые крики, по крайней мере у 4 исследованных форм фазана, хорошо отличаются на спектрограммах (рис. 1). Основные отличия - в расположении частотных энергетических максимумов сигналов и в длительности интервала между слогами крика. В дальнейшем это можно использовать в фазановодстве для диагностики подвида обыкновенного фазана.
Комфортные сигналы характерны как для самцов, так и для самок. У самцов сигнал звучит, как быстрое "ко-ко-ко-ко-кооо", причем количество слогов "ко" варьирует в зависимости от степени комфорта. Это самый низкочастотный сигнал из всего "акустического" репертуара фазана (0,2-0,6 кГц). Самки издают более высокое "коканье" (0,4-1,0 кГц). Характерная особенность сигналов комфорта - их ритмическая организация (рис. 2а, б), а такжe повышенная полифункциональность (издаются при отдыхе, насыщении, после копуляции и в других комфортных ситуациях). Очевидно эти сигналы могут быть использованы для снижения беспокойства птиц при стрессовых ситуациях (отлов, передержка и т.п).
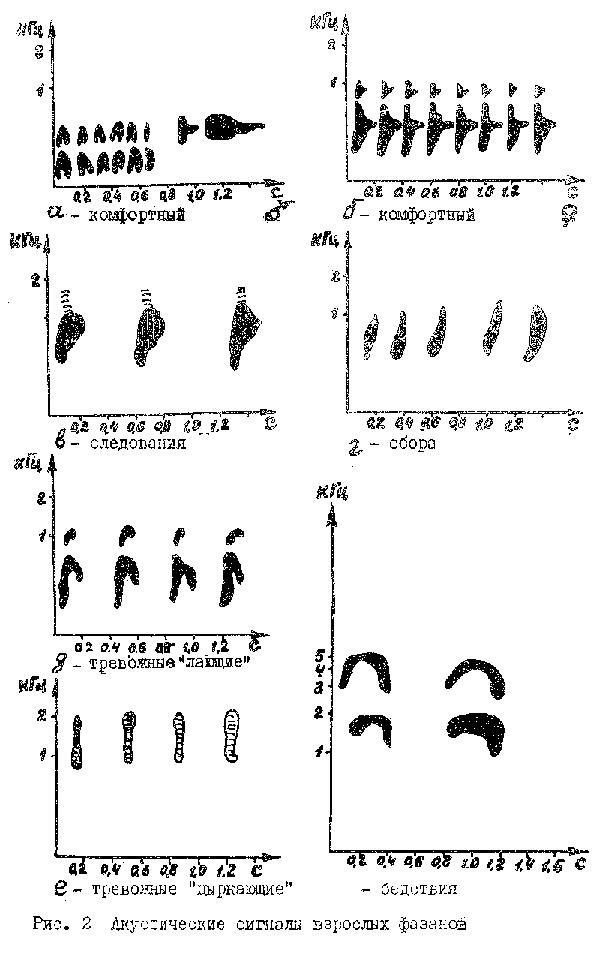
Сигнал взлета характерен только для взлетающих петухов и представляет собой трудно передаваемый слогами набор отрывистых повторяющихся звуков "ик-кирик-кирик-кирик". Этот сигнал можно рассматривать как тревожный.
Из других звуков необходимо выделить сигнал "бедствия", характерный в основном для самок, схваченных хищником или резко взятых в руки человеком. Эти же звуки издают и самки, не желающие спариваться с самцом при преследовании их последним. Характерной особенностью сигнала является высокая частота (3,5-6,0 кГц) и наличие частотной составляющей (рис. 2в). На слух сигнал "бедствия" воспринимается как тонкие пронзительные писки. Демонстрация записей этих сигналов фазанам, содержащимся в дичепитомнике, приводило к повышению их беспокойства (взлеты, побежки).
Звуковая сигнализация самок в родительский период
Характерно, что звуковые сигналы самки, адресованные птенцам, появляются еще до вылупления птенцев, за 1-2 дня до проклева скорлупы. В это время эмбрионы переходят от аллантоисного дыхания к легочному, в результате чего у них появляются первые звуковые сигналы: "щелкающие" звуки, сигналы "комфорта", "дискомфорта" (Тихонов, 1977). У самок фазана нами обнаружены следующие акустические сигналы.
Сигналы следования. Издаются самкой, уводящей птенцов от опасности или при дружном перемещении выводка. Сигнал звучит как мурлыкающее "трр-трр-трр". Каждая трелька состоит из 5-7 импульсов общей длительностью 150-250 мс.Частотная модуляция имеет форму зеркальной запятой (рис. 2г). Трельки объединяются в серии из 2-5трелек. Сигналы несут функцию оповещения о месте нахождения самки, призыва к следованию, информацию об опасности.
Сигналы сбора. У фазанов, очевидно, как и у кур (Отрыганьева, 1976; Тихонов, Отрыганьева, 1976), существует несколько сигналов сбора. Нами отмечен лишь 1 тип такого сигнала - резкие повторяющиеся (в ритме 3 импульса в секунду) звуки вроде "уить-уить-уить". Наиболее характерной чертой является резкое нарастание частоты сигнала (рис. 2д). Эти звуки, как и сигналы следования, можно использовать для сбора птенцов (Фокин, 1982б).
Тревожные сигналы. Начальная степень беспокойства (потенциальная опасность) характеризуется издаванием фазанкой "лающих" звуков, имеющих характерную форму частотной модуляции (рис. 2е). Приближение опасного объекта приводит к исполнению сигналов следования. Максимальная угроза сопровождается излучением высоких резких "цыркающих" звуков (рис. 2ж). Воспроизведение "цыркающих" звуков фазанятам вызывает у них бегство с последующим затаиванием, что можно использовать для специального обучения молодняка адекватно реагировать на тревожную ситуацию (Фокин, 1982а).
Акустические сигналы фазанят
Сигналы "дискомфорта" издаются фазанятами при охлаждении, голоде, изоляции от собратьев или от самки. Звуки представляют собой организованные в серии ритмически следующие друг за другом длительные (да 500 мс) писки. В каждой серии, как правило, 3-4 писка. Интенсивность их неоднозначна: первый издается тихо, он более короткий (100-200 мс), второй громче, третий и четвертый издаются с наибольшей интенсивностью и длительностью. Чем в большей степени птенец испытывает дискомфорт, тем интенсивнее и длительнее издаваемые звуки. Частотная модуляция преимущественно растянуто-колоколообразной формы (рис. 3а,б, в). Частотно-временные параметры сигналов приведены в табл. 1.
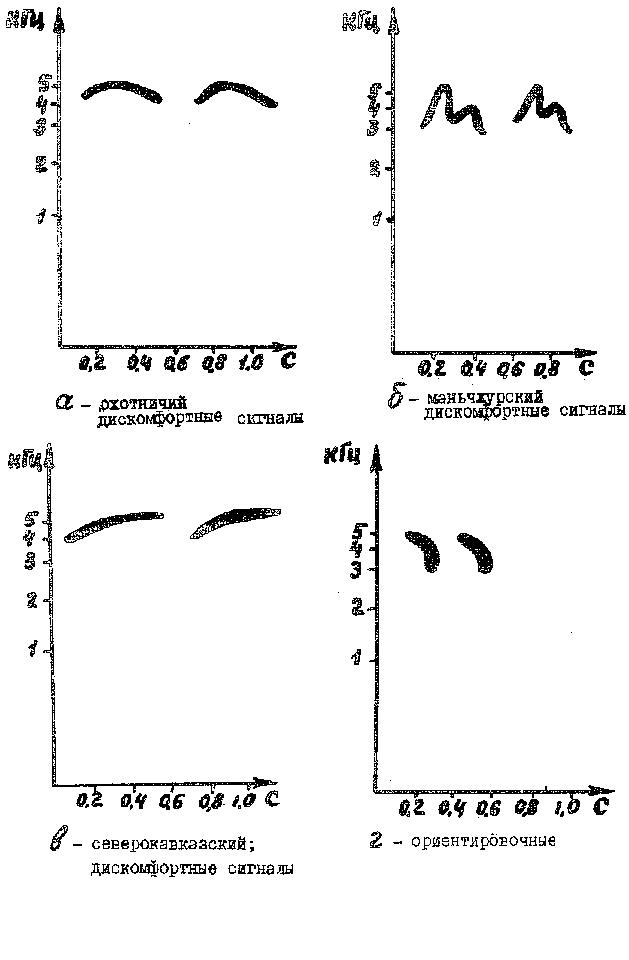
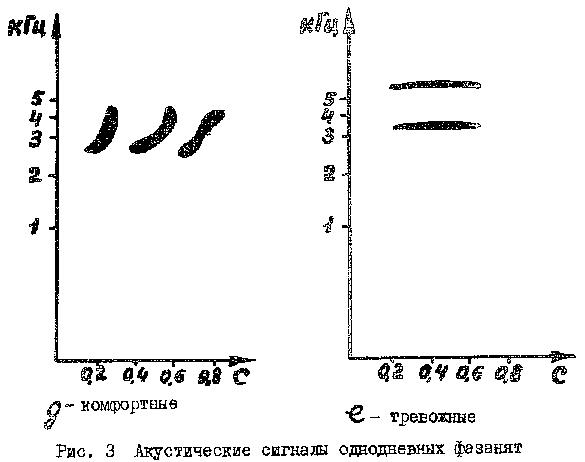
Таблица 1
Спектрально-временные характеристики акустических сигналов фазанят
|
Тип сигнала |
Спектральные границы (кГц) |
Длительность импульса (мс) |
Фронт нарастания интенсивности сигнала (мс) |
|
Дискомфортный |
2,9-6,5 |
40-300 |
|
|
Ориентировочный |
2,9-5,6 |
90-240 |
50-100 |
|
Комфортный |
2,6-5,1 |
25-180 |
20-110 |
|
Тревожный |
3,5-7,5 |
300-350 |
50-100 |
Во время излучения сигналов "дискомфорта" птенец вытягивает шею и издает звуки с открытым клювом. При охлаждении эти сигналы стимулируют взрослых птиц к обогреву птенцов.
Ориентировочные сигналы обычно предшествуют издаванию дискомфортных. Они издаются птенцами, попавшими в незнакомую обстановку или потерявшими контакт с выводком. Поведение птенцов в этих случаях характеризуется ориентировочной реакцией, в ходе которой фазаненок совершает быстрые повороты головы, прислушивается после каждой серии издаваемых им звуков. Затем следуют описанные выше сигналы "дискомфорта". Услышав аналогичные сигналы собратьев, птенец отвечает реакцией приближения, что обеспечивает интеграцию выводка. Физическая структура сигналов характеризуется организацией в посылки по 2-3 коротких импульса небольшой интенсивности (рис. 3г, табл. 1).
Комфортные сигналы птенцов наиболее многообразны (рис. 3д, табл. 1), причем функциональное значение различных вариантов не строго фиксировано. Это отмечено и у других выводковых птиц (Тихонов, Фокин, 1980). Однако можно выделить сигналы, характерные для ситуаций согревания птенцов (одиночные короткие писки), их объединения в группу (серии писков с плавно нарастающей частотой в каждой импульсе), насыщения и подготовки ко сну (трелевые звуки). Общий для всех комфортных сигналов является более низкая частота, меньшая длительность и небольшая интенсивность звучания. Сигналы издаются при закрытом клюве и снижении локомоторной активности.
Тревожные сигналы появляются у фазанят в условиях искусственного выращивания после 5-ти дневного возраста. Это одиночные, практически немодулированные звуки, издаваемые при беспокойстве птенцов людьми или при "стычках" между особями (рис. 3е).
Сигналы "бедствия" издаются птенцами в критических ситуациях (птенец пойман хищником, берется в руки за лапку и т. д.). В отличие от ряда других выводковых птиц, специализированные сигналы "бедствия" у фазанят отсутствуют. Например, у перепелят эти сигналы очень резко отличаются от других звуков. Фазанята же в этих ситуациях издают сигналы "дискомфорта", но с большей частотой следования звуков (3 писка в секунду вместо 1-2 при "дискомфорте"). В серии число писков может быть неограничено большим (при "дискомфорте" 2-4).
В практике дичеразведения возникла настоятельная необходимость разработки методов идентификация каждого из 12 подвидов обыкновенного фазана. Даже по экстерьерным признакам это удается с трудом и, в основном, только для самцов. Запись однодневных фазанят и последующий структурный анализ сигналов позволили нам сделать предположение о возможности определения по голосу принадлежности птенца к определенному подвиду. Анализу подвергались все типы акустических сигналов фазанят. Выяснилось, что различием обладают лишь сигналы "дискомфорта". Например, нижняя спектральная граница сигналов маньчжурского фазана лежит значительно ниже (2,9-3,5 кГц), чем у других исследованных форм (3,8-4,0 кГц). В то же время, длительность сигналов "дискомфорта" у него наименьшая (130-380 мс). У охотничьего фазана 240-400 мс, у северокавказского 260-600 мс. Основные различия наблюдались в форме частотной модуляции (рис. 3а,б,в). Это следует и из сонограмм, приводимых вработе В.Д. Ильичева и А.В. Тихонова (1979). Очевидно, форму частотной модуляции и следует положить в основу идентификации подвида фазанят.
В настоящее время проводятся и другие экспериментальные работы по фазану - по стимуляции с помощью звуковых сигналов пищевой активности фазанят, снижению их агрессивности, разрабатываются звуковые стимуляторы роста и развития молодняка, акустические методы повышения эффективности размножения. Применение биоакустических методов позволит не только сократить долю ручного труда в фазановодстве, но и обеспечит его более быстрое развитие в нашей стране.
Литература
Ильичёв В.Д., Тихонов A.В. Биологические основы управления поведением птиц. I. Куриные. - Зоол. журн., 1979, т. VIII, вып. 7, с. 1021-1031.
Отрыганьева А.Ф. Звуковые сигналы наседки и использование их для выборки цыплят из выводных лотков. - В кн.: Передовой опыт в птицеводстве. М., 1976, с. 3-7.
Тихонов А.В. Акустическая сигнализация и поведение выводковых птиц в раннем онтогенезе. - Автореф. канд. дис. М., 1977, 24 с.
Тихонов А.В., Отрыганьева А.Ф. Звуковая сигнализация у домашней курицы (наседки) и поведение цыплят. - Биологические науки, 1976, № 4, с. 77-82.
Тихонов А.В., Фокин С.Ю. Акустическая сигнализация и поведение куликов в раннем онтогенезе. II. Сигнализация и поведение птенцов. - Биологические науки, 1980, № 10, с. 45-54.
Фокин С.Ю. Биоакустика и дичеразведение. - Охота и охотн. хоз-во, 1982а, № 1, с. 4-5.
Фокин С.Ю. Использование акустической сигнализации птиц в искусственном дичеразведении.- В кн.: Разведение и создание новых популяций редких и ценных видов животных (тезисы докладов 3 совещания). Ашхабад, 1982б, с. 77-80.