




Франклин Я.Р. Эволюционные изменения в небольших популяциях // Биология охраны природы: Пер. с англ. / Под ред. М. Сулея, Б. Уилкокса. / Перевод Остроумова С.А.; Под ред. и с предисл. А.В. Яблокова. – М.: Мир, 1983. С. 160-176.
Аргументы в пользу охраны природы обычно основаны на экологических, экономических или даже этических соображениях. Однако имеются и важные генетические основания. Любая стратегия охраны конкретного вида должна, в частности, определяться знанием действительной или предполагаемой генетической структуры данного вида. Особенно строгие требования возникают, когда мы рассматриваем будущие потребности селекционеров растений и животных. Для того чтобы обеспечивать человечество продовольствием, а домашних животных — кормом, необходимо поддерживать разнообразие видов, причем каждый из них в достаточно большом количестве особей, чтобы иметь запас генетической изменчивости, адекватный потребностям селекционной работы. Потребности человека в этой области предсказать нелегко. Множество растений еще не изучалось как возможные ресурсы для получения масла, продуктов питания и волокна. Едва начаты исследования того, как можно использовать культуры клеток и тканей, включая культуры тканей и клеток животных. В настоящее время создаются банки семян (Harlan, 1975), имеются ограниченные запасы спермы некоторых домашних животных, но эта работа только начинается. Некоторые ценные сорта, породы и разновидности растений и животных мы уже потеряли. Находятся в опасности дикие родственники культурных сортов, служащие источником генетического материала для придания культурным сортам устойчивости к болезням и других полезных качеств.
Все эти вопросы важны, но я не хотел бы останавливаться на них. Данная глава будет посвящена обсуждению того, как малый размер популяций сказывается на эволюционных изменениях и выживании. Вопрос о сохранении генетических ресурсов уже подробно обсуждался (Frankel, 1974).
Мы знаем, что угрожаемый вид может поддерживаться в парках и резерватах в ограниченном числе особей. Из ограничения размеров популяции вытекает ряд генетических последствий. Возможно, наиболее важным из них, особенно в небольшие отрезки времени, является депрессия в результате инбридинга. Для многих видов она накладывает ограничение на минимальный размер популяции, необходимый для ее выживания. Однако даже в популяции, достаточно крупной для того, чтобы избежать серьезного инбридинга, может происходить постепенная утрата генетической изменчивости, что ограничивает дальнейшие эволюционные изменения. Я считаю, что популяция со случайно скрещивающимися особями, как у большинства млекопитающих и птиц, только для того чтобы избежать инбридинга, должна включать не менее 50 размножающихся особей. При рассмотрении более длительных отрезков времени генетическая изменчивость сохранится только при условии, что размер популяции будет на порядок выше. Последнее утверждение основано на следующих допущениях: 1) для выживания необходимы непрерывные и часто быстрые эволюционные изменения; 2) ответ на естественный отбор ограничен малым размером популяции.
Прежде чем детально обсудить эти положения, мы должны спросить, являются ли эволюционные изменения тем, чего мы хотим. Действительно, хотим ли мы сохранить современных слонов или обеспечить выживание их слоноподобных потомков? Это важный вопрос. В зависимости от того, чем мы озабочены: сохранением точного фенотипа вида или сохранением филогенетической линии, в которой мы допускаем непрерывные эволюционные изменения, — наша стратегия должна быть разной. В популяциях, разводимых в неволе, можно добиться максимальных эволюционных изменений, а в некоторых случаях, наоборот, почти полностью их затормозить.
При последующем обсуждении я буду иметь в виду в основном популяции со случайно скрещивающимися особями и филетическую эволюцию.
АДАПТАЦИЯ
Адаптация и вымирание
Исторические документы дают много примеров эпизодического вымирания и быстрых эволюционных изменений. Самые последние из них начались в позднем плейстоцене с исчезновением многих видов крупных млекопитающих. Относительно причин плейстоценовых вымираний есть противоречивые суждения. Некоторые приписывают их климатическим изменениям, другие - росту экологического доминирования человека. Однако в отношении причин современного кризиса сомнений нет. В дополнение к прямой конкуренции и хищничеству со стороны человека в мировой биоте происходит активная «перетасовка». В Австралии, например, в прямую конкуренцию с местной флорой и фауной вступают интродуцированные плацентарные млекопитающие, декоративные растения и пастбищные виды. Сверх всего этого происходит крупномасштабное разрушение местообитаний, которое хорошо документировано.
В ответ на это экологическое разрушение мы можем ожидать, и уже наблюдаем, бурный всплеск эволюционных изменений. В течение ряда лет мы отмечаем развитие индустриального меланизма, появление резистентных к ядохимикатам насекомых, иммунных к миксоматозу кроликов и трав, которые могут расти на отвалах свинцовых и медных рудников. Некоторые виды легко приспосабливаются к новой для них среде, другие, - в основном по неизвестным причинам, не могут приспособиться и оказываются под угрозой вымирания. Непосредственной причиной вымирания может быть случайное событие, такое, как пожар или эпидемия, но вид, уязвимый к этим случайным явлениям, по-видимому, таков потому, что он в прошлом хуже, чем его конкуренты, приспособился к меняющейся среде. В конечном счете, если популяция не может поддерживаться физической или биотической средой, она - исчезает. Адаптация — критически важное понятие, совершенно необходимое при любом анализе выживания популяции.
Адаптация и естественный отбор
Адаптация, приспособление или модификация тех черт, благодаря которым организм наиболее удачно «встраивается» в среду, представляет собой процесс, известный только по его последствиям. Результаты адаптации (например, структура зубов, приспособленных к подрезанию или ощипыванию, защитная окраска и мимикрия, миграционное и территориальное поведение, опыление и механизмы распространения семян) распознаются по связи между структурой и функцией. Реже мы понимаем адаптацию в терминах изменения частот генов, так как в целом мы еще далеки от того, чтобы иметь возможность переводить физиологию и поведение на язык частот появления и исчезновения известных генотипов.
Несмотря на неполноту наших знаний, можно предполагать, что наиболее важные адаптивные признаки — это непрерывно варьирующие (количественные) характеристики. Поскольку адаптация является результатом естественного отбора (хотя не любой естественный отбор адаптивен), мы можем привлечь значительный опыт работы по отбору в лаборатории и на домашних животных, для того чтобы сделать ряд заключений о связи размера популяции и эволюционных изменений.
Обычно различают 3 вида отбора количественных признаков: 1) стабилизирующий отбор, или отбор, направленный против экстремальных значений признаков; 2) направленный отбор, при котором только одно крайнее значение признака имеет преимущество; 3) дизруптивный отбор, или отбор в пользу всех экстремальных значении признаков и направленный против промежуточных значений признаков.
Нет сомнения, что стабилизирующий отбор преобладает в природных популяциях. Для многих признаков наибольшая приспособленность (измеряемая по выживанию или числу потомков) характерна для особей со значениями данных признаков, близкими к средним по всей популяции. Примером могут служить размер раковины у моллюсков (Weldon, 1901), размер яйца у уток (Rendel, 1943), размер кладки у стрижей (Lack, 1954), высота взрослых и вес новорожденных у человека (Cavalli-Sforza, Bodmer, 1971). Вместе с тем, направленный отбор, будучи обычным при селекции животных и растений, в природе, по-видимому, редок. Полагают, что наиболее приспособленные особи «проявляют гармоничное сочетание всех признаков» (Lerner, 1954), в то время как экстремальные особи, которые «приспособлены» по одному из признаков, оказываются не сбалансированными в отношении других. Следовательно, при направленном отборе по одному признаку приспособленность часто уменьшается.
Третий тип отбора— дизруптивный — недостаточно хорошо изучен. Суждения о его важности в природных популяциях противоречивы. Вероятно, он действует в среде с пятнистыми местообитаниями и может играть роль в видообразовании.
Итак, естественный отбор — это довольно сложный процесс, в котором медленное изменение видов сопровождается медленными изменениями ниши. Вид «всегда остается чуть-чуть позади, немного не достигая идеальной приспособленности; в конце концов, если вид отстает от изменений среды из-за недостаточной генетической изменчивости, на основе которой работает естественный отбор, он становится вымирающим (Lewontin, 1978). Поскольку при отборе меняется каждый признак, скорость приспособительного ответа есть функция интенсивности отбора и наследуемости признака. Оба этих качества подвергаются воздействию генетического дрейфа. Хорошо известно, что популяции, которые «прошли через бутылочное горлышко» или поддерживаются при малой численности, не проявляют такого сильного ответа на искусственный отбор, как популяции с большим числом особей.
ГЕНЕТИЧЕСКИЙ ДРЕЙФ И ЭФФЕКТИВНЫЙ РАЗМЕР ПОПУЛЯЦИИ
В конечной популяции сочетание генотипов в каждом поколении формируется выборкой гамет из предыдущего поколения. Почти все генетические эффекты, которые возникают в малых популяциях, являются результатом нерепрезентативности выборок. Этот процесс называется генетическим дрейфом. В малых популяциях частоты генов изменяются от одного поколения к другому даже в отсутствие отбора, мутаций или миграций. Эти случайные изменения ведут к постепенному росту гомозиготности. Если они ничем не ограничиваются, то приводят к фиксации определенных аллелей во всех локусах и, следовательно, к полной потере генетической изменчивости. На уровне фенотипа мы можем часто наблюдать следующие 3 эффекта генетического дрейфа: 1) инбредную депрессию в результате либо потери гетерозиготности, либо фиксации неблагоприятных генов; 2) случайные изменения фенотипа, особенно изменения средних значений количественных признаков; 3) снижение генетической дисперсии (и, следовательно, понижение наследуемости).
Для популяций данного размера степень генетического дрейфа варьирует в зависимости от состава группы размножающихся особей и от распределения числа потомков. Поэтому теоретические последствия дрейфа обычно рассчитывают для идеальной популяции, в которой каждая особь дает равный вклад гамет в общий фонд, из которого формируется новое поколение.
Каждая реальная популяция из N особей имеет выборочную дисперсию в частотах генов, которая эквивалентна выборочной дисперсии идеальной популяции, состоящей из Neособей. Таким образом, Neесть эффективный размер популяции, которая действительно состоит из N особей. Это важная концепция. При дальнейшем обсуждении инбредной депрессии или потери генетической дисперсии будет чаще использоваться именно эффективная численность* (или эффективный размер популяции), а не численность, выявляемая при учете (реальная численность).
* Примечание к электронной версии: «эффективная численность популяции» - количество особей в популяции, принимающих участие в размножении. В. Гагарин.
Кимура и Кроу (Kimura, Crow, 1963) дали исчерпывающее изложение этой теории. Мы проиллюстрируем здесь связь между действительной и эффективной численностью популяции с помощью нескольких важных примеров.
1. Эффект варьирования числа потомков.Пусть дисперсия числа потомков будет ?2, а действительная численность популяции — N. Тогда Ne = 4N/2+?2 (приблизительно). Допустим,например, что средняя численность семьи равна 2, а дисперсия равна 4. Тогда Ne=(2/3)N. Вместе с тем, если каждая размножающаяся пара дает следующему поколению ровно двух потомков, т. е. численность семьи не варьирует, то Ne = 2N. Эффективный размер популяции снижается при росте варьирования в числе потомков, и, наоборот, Neстановится максимальным, когда все семьи дают в следующее поколение равные вклады. Исходя из наблюдавшихся распределений числа потомков у разных видов, подсчитали, что эффективный размерпопуляции варьирует от 60 до 85% реального числа особей в популяции (Crow, Morton, 1955).
2. Неравное количество особей разного пола.Если число самцов, дающих гаметы для следующего поколения, и число самок различается и Nm— число самцов, a Nf— число самок, то
1/ Ne= ?Nm + ? Nf(1)
Это особенно важно для видов, которые имеют постоянные гаремы (например, слоны, тюлени, зебры, викуньи и летучие мыши). Размножающаяся популяция, в которой имеется 90 активных самок и лишь 10 активных самцов, имеет эффективный размер не 100, а 36.
3. Флуктуации численности популяции.Если численность популяции варьирует от поколения к поколению, то эффективный размер есть, гармоническая средняя:
1/ Ne=1/t(1/ N1+1/ N1+ . . . +1/ Nt) (2)
где Ntесть эффективная численность t-гопоколения. Например, допустим, что популяция с обычным эффективным размером 1000 уменьшается в одном из поколений до 50. Тогда в интервале десяти поколений:
Ne=10/(1/50+9/1000)=345 (3)
На эффективный размер оказывают существенное воздействие даже редко происходящие снижения численности, возможные при засухе или болезни.
При рассмотренном подходе к эффективному размеру допускается, что каждая особь случайным образом образует пару для размножения с другими членами данной группы. Такое допущение может быть верным для видов, включающих очень подвижных особей, но оно неадекватно для многих растений и нелетающих животных. У последних инбридинг — обычное явление; популяция часто резко подразделяется на много мелких групп, между которыми происходит инбридинг. В случаях, когда степень связи между активными в плане размножения особями есть функция их географического удаления друг от друга, генетический анализ структуры популяции значительно усложняется.
КРАТКОВРЕМЕННЫЕ ЭФФЕКТЫ: ИНБРЕДНАЯ ДЕПРЕССИЯ
Этот вопрос здесь будет затронут лишь кратко. Эмпирически ясно, что виды значительно различаются по своему отношению к инбредной депрессии. Многие растения в природе размножаются с помощью самоопыления. Среди животных наиболее чувствительны к инбредной депрессии млекопитающие и птицы, вероятно, потому, что для них характерна низкая скорость размножения. Именно в этом случае инбредная депрессия оказывается наиболее серьезным последствием уменьшенного размера популяции.
Степень инбридинга в популяции измеряют с помощью коэффициента инбридинга f. Если эффективный размер популяции Ne, то за каждое поколение f увеличивается на ?Ne. Таким образом, в малых популяциях постепенно, за многие поколения, инбредная депрессия накапливается. Даже в случае тесного инбридинга при образовании популяции, берущей начало от небольшого числа особей, потомки не обязательно окажутся сильно пострадавшими от инбредной депрессии. Уровень инбридинга легко снижается при иммиграции в такую популяцию неродственных особей.
Инбридинг оказывает разрушительное воздействие на выживание и репродукцию. Он влияет на такие характеристики, как скорость роста и размер взрослых особей. Для многих признаков инбредная депрессия в долях среднего значения составляет около трети от коэффициента инбридинга. Для плодовитости она, возможно, вдвое больше. В целом воздействие инбридинга на приспособленность, вероятно, значительно выше. На дрозофиле показано (Latter, Robertson, 1962), что влияние инбридинга на конкурентный индекс (что для наших целей равносильно приспособленности) описывается выражением е-2,7f. Это значит, что для малых fснижение приспособленности в 2,7 раза превышает коэффициент инбридинга. В случае гомозиготности Drosophilapseudoobscuraпо одной из аутосом, составляющей около одной пятой генома, приспособленность снижалась почти на 20% (Sved, Ayala, 1970). Несмотря на эти разрушительные эффекты, с небольшой степенью инбридинга все же можно мириться. Животноводы допускают у домашних животных коэффициент инбридинга до 1% на поколение (т. е. Ne = 50). Естественный отбор, конечно, будет учитывать быстрые отрицательные эффекты генетического дрейфа. Для каждого вида имеется своя минимальная численность популяции, при которой популяция способна справиться с эффектами инбридинга хотя бы за счет снижения выживаемости и скорости размножения. По-видимому, в отсутствие регулярной интродукции неродственных особей минимальный эффективный размер популяции для крупных млекопитающих составляет, по крайней мере, 50.
Наконец, важно подчеркнуть, что степень инбридинга увеличивается со скоростью, которая прямо связана с длительностью жизни одного поколения. Популяция из 50 слонов, случайно образующих пары, достигнет 10%-ой степени инбридинга через несколько сот лет. Та же степень у мышей будет достигнута менее чем через 10 лет.
ДОЛГОВРЕМЕННЫЕ ЭФФЕКТЫ
Даже в популяциях, где нет инбредной депрессии, эффект выборки вызывает флуктуации частот генов которые имеют важное значение для будущей эволюции вида. Некоторые гены будут случайно утеряны — особенно исходно редкие в популяции. Частота аллелей, поддерживаемая с помощью отбора на некотором промежуточном уровне, впоследствии отклоняется от равновесия. Теория отбора в ограниченных популяциях достаточно сложна. Следует отметить, что результаты сильно зависят от допущений, сделанных относительно типа естественного отбора.
На фенотипическом уровне мы наблюдаем изменения в частотах просто наследуемых признаков. Для количественных признаков можно наблюдать дрейф в среднем значении и чаще всего - снижение генетического разнообразия. Последние два эффекта мы обсудим более детально, причем хотелось бы отметить, что критическое значение имеет именно утрата генетического разнообразия.
Случайные изменения фенотипа
У линий лабораторных животных, подвергшихся инбридингу, помимо общих эффектов инбредной депрессии наблюдаются изменения фенотипа. Например, отмечалось (Wright, 1977a), что линии морских свинок, подвергшихся инбридингу, различаются по морфологии, физиологии, окраске и темпераменту. В анализе эволюционных процессов Райт придает большое значение сдвигам средних значений признака в малых популяциях, поскольку только путем генетического дрейфа создаются новые адаптивные комбинации признаков. Следовательно, предполагает Райт, расщепление вида на изолированные субпопуляции облегчает эволюционные изменения. Эта широко распространенная точка зрения отличается от позиции Фишера, который считает, что эволюционные изменения максимальны в больших популяциях.
Поскольку для большинства количественных признаков естественный отбор обычно благоприятствует промежуточным значениям, любое изменение уже хорошо адаптированного организма будет разрушительным и столкнется с противодействием естественного отбора. Теоретически можно показать, что дрейф в популяциях достаточного размера (например, из 500 особей) невелик и естественный отбор вызовет лишь небольшую смертность. На деле наблюдаемые изменения средних значений признаков в малых популяциях, вероятно, указывают на незначительную адаптивную важность таких изменений в данных условиях.
Истощение генетической дисперсии
Наблюдаемая фенотипическая дисперсия может быть разделена на три компонента; дисперсия, обусловленная изменениями среды, генотипическая дисперсия и дисперсия, обусловленная взаимодействием генотипа и среды. Последний компонент часто недооценивают. Он важен, поскольку генотип, наилучший в определенной среде, не обязательно окажется наилучшим в другой. В генотипической дисперсии можно выделить аддитивную, доминантную и эпистатическую составляющие. Популяция немедленно реагирует на естественный отбор именно с помощью аддитивной дисперсии (иногда ее называют генетической или генной дисперсией). Отношение аддитивной дисперсии к фенотипической называют наследуемостью. Если наследуемость признака равна нулю, то адаптивные изменения маловероятны, если вообще возможны.
Наше понимание сил, лежащих в основе поддержания генетической дисперсии количественных признаков, крайне поверхностно. Однако мы знаем, что разнообразие повсеместно и что большинство признаков с готовностью откликнется на отбор. Первоначальным источником генетической изменчивости являются, конечно, мутации. Но вероятность того, что новая мутация достигнет разумной частоты, зависит от скорости мутирования, численности популяции и типа отбора, в данный момент действующего на признак. При направленном отборе любая мутация, которая благоприятствует изменению, но неразрушительна в других отношениях, будет способствовать увеличению частоты. Однако стабилизирующий отбор позволит накапливаться лишь генам со слабым действием (Fisher, 1958).
Рассмотрим только скорости приобретения и утраты аллелей при мутировании и генетическом дрейфе, пренебрегая эффектами, связанными с естественным отбором. В очень малых популяциях утрата изменчивости при взятии выборки будет больше, чем ее приобретение в результате мутирования, и таким образом в итоге будет наблюдаться уменьшение изменчивости. Наоборот, в очень больших популяциях влияние мутационного процесса будет доминировать, и можно ожидать устойчивого увеличения изменчивости. Для каждого признака существует своя численность популяции, при которой скорости приобретения и утраты изменчивости уравниваются, т. е. в. данном случае ее уровень уже не будет больше меняться. Если доминантная или эпистатическая дисперсии отсутствуют, то аддитивная дисперсия утрачивается с той же скоростью, что и гетерозиготность, т. е. со скоростью ? Ne за поколение. Эта цифра, по-видимому, достаточно точна для большинства признаков. Скорость приобретения изменчивости при мутировании определить сложнее. У нас есть некоторая скудная информация о скорости возникновения новых вариаций для такого признака, как число щетинок в гомозиготных линиях дрозофил. По данным, обзор которых сделал Ланде (Landе, 1976), эта скорость составляет около одной тысячной от дисперсии, обусловленной изменениями среды. Поэтому для числа щетинок мы ожидаем примерное равновесие между приобретением и утратой изменчивости при ?Ne = 10-3 что позволяет определить Ne = 500. Отбор и сцепление генов значительно осложняют эту простую картину, но я попытаюсь показать, что даже сильный отбор не приводит к резкому снижению аддитивной дисперсии.
Существующий уровень генетического разнообразия в популяции в большой степени определяется предыдущим отбором (за время, прошедшее с последнего резкого сокращения численности популяции) и эффективным размером популяции. Влияние продолжающегося стабилизирующего отбора оценить очень трудно, но по некоторым данным аддитивная дисперсия подвергается существенному воздействию. Согласно теории, стабилизирующий отбор снижает все компоненты фенотипической дисперсии. Уменьшение генотипического компонента наблюдается вследствие фиксации генов, роста нарушений равновесия в результате эффекта сцепления (Bulmer, 1976) и в конечном счете развития онтогенетических систем обратной связи, которые эволюционируют в направлении стабилизации фенотипа (Waddington, 1957). Однако это очень медленные процессы. Было показано (Kimura, 1965; Latter, 1970; Lande, 1976), что существенная генетическая дисперсия может поддерживаться мутационным процессом при стабилизирующем отборе.
Направленный отбор также снижает аддитивную дисперсию, но опять же медленно. Теоретически скорость изменений генетической дисперсии пропорциональна третьему моменту генных эффектов. Если, как подсказывает здравый смысл, количественные признаки контролируются большим числом генов (каждый с малым эффектом), то третий момент должен быть близок к нулю и генетическая дисперсия будет меняться очень медленно. В добавление к этому не исключено, что тесно сцепленные гены действуют как хранилище изменчивости. Под давлением отбора эта изменчивость может медленно «высвобождаться» посредством рекомбинаций.
В основном экспериментальные данные подтверждают теоретические аргументы. Снижения дисперсии под действием стабилизирующего отбора достигнуть трудно, но ряд авторовполучили положительные результаты (Rendel, 1960; Prout, 1962). Однако их эксперименты трудно интерпретировать; не всегда ясно, какой именно компонент дисперсии (генетический или обусловленный изменениями среды) изменился. Более понятны последствия направленного отбора, которые анализировали в многочисленных экспериментах. Один из наиболее длительных экспериментов был проведен в штате Иллинойс, где наблюдали реакцию кукурузы на отбор по содержанию масла и белка в течение 76 поколений. Отбор был начат в1896 г. с исходной популяции из 163 открыто опыляемых початков. В каждом поколении отбирали 12-24 экземпляров родительских растений. Реагирование на отбор по содержанию масла показано на рис. 1; сходная закономерность наблюдалась и в случае отбора на содержание белка. В течение экспериментабыл достигнут ответ, составляющий 20 стандартных отклонений аддитивной дисперсии в сторону увеличения содержания масла и белка. Менее эффектные результаты были получены для отбора (в довольно больших популяциях) у различных лабораторных животных, таких, как Drosophila, Triboliumи мышь. Примеры можно найти в работе Фалконера (Falconer, 1960). Проведенные эксперименты убедительно показывают, что аддитивная генетическая дисперсия истощается отбором не очень быстро.
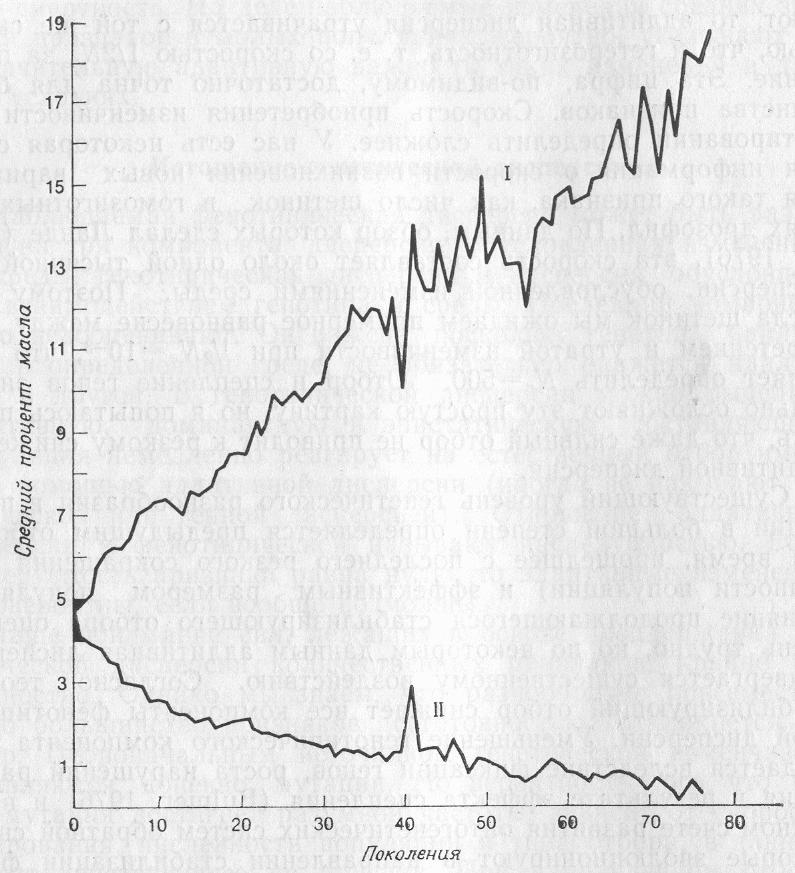
Рис. 1. Средний процент масла в початках кукурузы Zeamaysпри отборе на его высокое (I) и низкое (I) содержание (Dudley, 1977)
Какой же вывод мы можем сделать о действии естественного отбора на генетическую дисперсию? Сильный отбор уменьшает аддитивную дисперсию, но слабый направленный - или стабилизирующий отбор, вероятно, оказывает малое воздействие. В некоторых опытах даже при длительном отборе в изучаемой популяции была такая же аддитивная дисперсия, как и в исходной. Поэтому в порядке рабочей гипотезы мы можемзаключить, что основным фактором, определяющим уровень генетической дисперсии в природных популяциях, являетсяравновесие между генетическим дрейфом и мутациями.
ЭФФЕКТ ОСНОВАТЕЛЯ И РЕАГИРОВАНИЕ НА ОТБОР
В предыдущем разделе были рассмотрены длительные эффекты малого размера популяции. Здесь я хочу проанализировать эффект «бутылочного горлышка» (резкого увеличения численности популяции после того, как устраняются факторы, ограничивающие ее рост), или основания популяции группой из малого числа особей. Экспериментальные исследования в малых популяциях (Frankham et al., 1968; Hanrahan et at, 1973) показывают, что ответ на отбор в них меньше, чем в крупных популяциях, но в основном эта разница не очень велика. Так, даже в наиболее экстремальных ситуациях, когда популяция была основана единственной парой особей, сохраняется при четверти аддитивной дисперсии. Это означает, что в среднем сохраняется возможность для заметного реагирования.
Хэммонд (Hammond, 1973) собрал данные, которые хорошо иллюстрируют эффект небольшого числа основателей популяции. Он создал популяции дрозофилы на основании одной, десяти и пятидесяти пар и затем измерил реагирование на отбор по абдоминальным щетинкам в течение 10 поколений. Эти данные (табл. 1) показывают, что реализованные наследуемости (мера наблюдаемого изменения) в популяциях, созданных на основе 10 пар и на основе 50 пар, почти не различаются. Как и ожидалось, популяции, основанные одной парой особей, но поддерживаемые при более высокой численности, проявляют три четверти максимального ответа.
Таблица 1
Зависимость между численностью первоначальной популяции и последующей реакцией на отбор (Hammond, 1973)
|
Численность первоначальной популяции |
Режим регулирования популяции1 |
Число повторностей |
Реализованная наследуемость |
|
1 пара |
1-1 |
28 |
0,051 + 0,018 |
|
1-10 |
21 |
0,157 + 0,007 |
|
|
1-20 |
18 |
0,152 + 0,007 |
|
|
10 пар |
10-10 |
24 |
0,185 + 0,004 |
|
10-20 |
21 |
0,177 + 0,006 |
|
|
50 пар |
50-50 |
17 |
0,202 + 0,006 |
|
50-100 |
9 |
0,205 + 0,006 |
1 В данном столбце первая цифра — это число пар особей, до которого была снижена численность популяции в одном поколении, а вторая — это число пар, которое поддерживалось в последующих поколениях. Например, 1-10 означает, что численность популяции в первом поколении была равна одной паре особей, а затем популяция поддерживалась при численности 10 пар.
Хотя эффекты основателей и не могут существенно ограничивать немедленный ответ на отбор, тем не менее, они приводят к большим изменениям генофонда, так что характер наблюдаемых изменений может сильно отличаться от характера изменений в той популяции, из которой берет начало данный генофонд. Важный теоретический вклад в теорию «бутылочного горлышка» сделали Ней и др. (Nei еt al., 1975). Создание новых популяций на основе малого числа особей имеет существенное эволюционное значение. Этот вопрос заслуживает значительно большего внимания со стороны теоретиков и экспериментаторов, чем то, которое уделялось ему до сих пор.
УПРАВЛЕНИЕ И ОХРАНА УГРОЖАЕМЫХ ВИДОВ
До сих пор я рассматривал влияние малой численности популяции на ее эволюционный потенциал при условии, что вмешательство со стороны человека отсутствует. Теперь я хочу бегло рассмотреть последствия такого вмешательства. В некоторых условиях, в частности, когда размножающаяся популяция поддерживается в зоопарке или резервате, те решения, которые связаны с размножением, имеют эволюционные последствия. Следует помнить, что программы, в которых предполагается свести к минимуму инбридинг и генетический дрейф, не благоприятствуют одновременным эволюционным изменениям.
Например, один из путей поддерживать эффективный размер популяции — сделать так, чтобы каждая семья давала одинаковый вклад в следующее поколение в результате выбраковки особей только внутри семей. Существует и другая возможность (если она реальна), а именно таким образом подбирать пары для размножения, чтобы максимально избежать инбридинга. Все такие методы снижают ответ на естественный отбор и замедляют элиминацию особенно ненормальных генотипов. Однако если есть возможность выбора, то, по-видимому, лучше в данный момент обойтись без эволюционных изменений чтобы сохранить генетическое разнообразие для будущих эволюционных изменений. Это особенно важно в случае разведения в неволе, когда предполагается выпуск животных в дикую природу. В таких случаях следует избегать доместикации.
Хотя доместикация не заменяет сохранение генофонда в полном смысле слова (известно, что она необратима), по-видимому, в будущем в целях сохранения вида с доместикацией придется мириться. Я не хочу углубляться в эту область, отмечу только, что многие из поведенческих особенностей, которые сопровождают доместикацию, по-видимому, имеют высокую наследуемость и могут быть селектированы относительно легко (Fuller, 1969). Об изменениях, которые наблюдаются в процессе доместикации, известно очень мало. Нам необходимо идентифицировать важнейшие признаки, изменяющиеся при доместикации, понять их наследуемость и, возможно, каталогизировать те виды, которые предрасположены к комменсализму с человеком. Прекрасный обзор эволюции доместикации сделал Хейл (Hale, 1969).
Наконец, необходимо решить, что целесообразнее: поддерживать единственную большую популяцию или разбить данный вид на несколько небольших размножающихся групп. При этом прежде всего принимаются во внимание соображения политики или экологии, но, по-видимому, вариант нескольких небольших популяций имеет отчетливые генетические преимущества. В этом случае не только снижается опасность случайного вымирания (например, в результате заболевания), но создается также основа для локальных адаптаций, что увеличивает вероятность окончательного выживания. Генетический дрейф может быть уравновешен допущением случайных миграций. Мы знаем, что в отсутствие отбора субпопуляции имели бы свойства единой группы (особи которой скрещиваются друг с другом), если бы в течение каждого поколения происходил обмен одной-двумя мигрирующими особями. Относительно результатов совместного действия отбора и миграции в малых подразделенных популяциях информации очень мало. Однако теоретический анализ (Avery, 1978) позволяет предположить, что скорости миграции порядка одной особи на поколение увеличивают общую генетическую изменчивость идля гетерозисных локусов, и для локусов, которые подвергаются в субпопуляциях отбору в противоположных направлениях.
РЕЗЮМЕ
О силах отбора, поддерживающих полиморфизм по отдельным генам или адаптированным генным комплексам, мы знаем очень мало; я не рассматривал последствия генетического дрейфа на этом уровне. Вместо этого я анализировал проблему сохранения генетической изменчивости с точки зрения количественной генетики. В последней уделяется больше внимания средним значениям и дисперсиям различных непрерывно изменяющихся признаков, чем изменениям частот гена, средней гетерозиготности или долям полиморфных локусов. Основываясь на современных знаниях о количественных признаках, я пришел к предположению,что эволюционный потенциал популяций, поддерживаемых при достаточных эффективных численностях (порядка сотен, но не десятков особей), не находится под серьезной угрозой. Я думаю, что большинство количественных генетиков, работающих с популяциями животных, пришли бы к тому же выводу.
Объект изучения количественной генетики — не столько природные популяции, сколько организмы, размножающиеся в лаборатории, или домашние животные. Наиболее важные адаптивные изменения, которые наблюдаются в процессе эволюции, являются изменениями непрерывно меняющихся признаков. Поэтому есть все основания считать, что теория отдельных генов неадекватна для нашего понимания динамики таких изменений. Экстраполяция от лабораторных к природным популяциям резко осложняется различиями двух типов окружающей популяцию среды. Хорошо известно, что уменьшение численности или исчезновение одного вида оказывает воздействие и на другие виды.
Следует признать, что наше понимание рассматриваемых межвидовых отношений неполно. Недостаточно также изучен важный вопрос об отборе, зависящем от плотности. Признак, который благоприятен при высокой плотности, может быть вредным при низкой. Следовательно, снижение численности популяций способствовало бы увеличению генетического груза. Таким образом, любое изменение в экосистеме потенциально способно влиять на характер адаптации, и поэтому вид может оказаться в состоянии серьезного кризиса, если он столкнется сразу со слишком многими изменениями. Негативные генетические корреляции усугубляют ситуацию, ибо если мы пытаемся вести отбор сразу по двум антагонистическим признакам, то вряд ли можем ожидать существенных изменений, хотя бы одного из них.
Я уже отмечал, что в кратковременном варианте эффективная численность популяции должна быть не менее чем 50 особей. Как рабочую гипотезу я выдвигаю предположение, что для более длительного существования минимальная эффективная численность популяции должна быть 500. При более низких значениях численности увеличивается вероятность того, что генетическая дисперсия сложного признака будет утрачиваться существенно скорее, чем восстанавливаться с помощью мутаций. При разработке этого заключения я принял точку зрения, что важно поддерживать фонд разнообразия, на основе которого в будущем сможет действовать отбор. Число, к которому я пришел, основано на крайне скудных данных, и я подчеркиваю: для того чтобы ответить на обсуждаемые вопросы с достаточной уверенностью, необходимы гораздо более подробные исследования. Нет сомнения, что вид, эффективная численность которого в десять раз больше этого значения, будет находиться под меньшей угрозой генетического вырождения.
Некоторые виды способны восстанавливаться из очень малых популяций. Так, у морского слона численность одно время резко снизилась, и он, по-видимому, утратил значительную часть своей гетерозиготности (Bonnel, Selander, 1974). Тем не менее, его способность реагировать на отбор неизвестна. Другие виды могут оказаться под угрозой и при большой эффективной численности популяции. Популяции с зависимым от плотности темпом воспроизводства могут иметь критическую численность (значительно выше 500), нижекоторой популяция обречена на вымирание по чисто экологическим причинам.
Хотя я считаю, что снижение численности незначительно лимитирует реакцию на отбор, это не означает, что популяция всегда способна адаптироваться к новым условиям. Рост генетического груза вследствие изменений в экосистеме может быть столь значительным, что необходимые для адаптации генетические изменения — будь численность популяции 10 особей или 10 млн. особей — окажутся невозможными при репродуктивной способности данного вида.
Наконец, следует указать, что рассматриваемые здесь выводы касаются эволюционных изменений в легко идентифицируемых, случайно скрещивающихся субпопуляциях, поэтому они больше относятся к видам высокоподвижных животных. С точки зрения сохранения генетического разнообразия вида один слон может быть так же хорош, как и другой. Однако такой подход, безусловно, неправилен для большинства растений, где надо учитывать микродифференциацию иварьирование экотипов. Если вы видели одну секвойю, то это еще не значит, что все остальные такие же.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
Количественная генетика
Falconer D. 1960. Introduction to Quantitative Genetics, Oliver and Boyd, London.
Pollak E.О., Kempthome О., Bailey T. (eds.). 1977. Quantitative Genetics, Iowa State University Press, Ames, Iowa. Сборникстатей в данной области, который содержит несколько очень полезных обобщающих работ.
Hilt W. G. 1972. Estimation of genetic change (I. General theory and design of control poрulations), Animal Breeding Abstracts, 40, 1-15. Этот отличный обзор по проектированию контролирования популяций имеет прямое отношение к управлению видами в зоопарках и природных резерватах.
Охрана генетических ресурсов
Frankel О.Н. 1974. Genetic conservation: оur evolutionary responsibility Genetics, 78, pp. 53-65.
Harlan J.R, 1975. Our vanishing genetic resources, Science, 188, pp. 618-621.
Brown A.H.D. 1978. Isozymes, plant populations, genetic structure and genetic conservation, Theoret. Appl. Genet., 52, pp. 147-157.Статьи Хэрлана и Брауна представляют собой обзор и обсуждение проблем, связанных с растительными ресурсами.
Сентябрьский выпуск журнала Scientific American за 1978 г. полностью посвящен эволюции. Статья Левонтина в этом выпуске журнала имеет прямое отношение к проблемам, рассматриваемым в данной главе. [Вся подборка статей по эволюции из этого журнала переведена на русский языки и выпущена отдельной книгой («Эволюция», М.: Мир, 1981). Прим. перев.]
ЛИТЕРАТУРА
Avery P.J., 1978. The effects of finite population size on models of linked overdominant loci, Genet. Res. Camb., 31, 239-254.
Bonnel M.L, Selander R.K., 1974. Elephant seals: genetic variation and near extinction, Science, 184, 908-909.
Bulmer M.G., 1976. The effect of selection on genetic variability: A simulation study, Genet. Res. Camb., 28, 101-117.
Cavalli-Sforza L.L., Bodmer W.F., 1971. The Genetics of Human Populations, W.H. Freeman and Company, San Francisco.
Crow J.F., Morton N.E., 1955. Measurement of gene frequency drift in small
populations, Evolution, 9, 202-214.
Falconer D.S., 1960. Introduction to Quantitative Genetics, Oliver and Boyd Ltd., London.
Fisher R.A.,1958. The Genetical Theory of Natural Selection, 2nd ed., Dover
Publications, Inc., New York.
Frankel О.Н., 1974. Genetic conservation; our evolutionary responsibility, Genetics, 78, 53-65.
Frankham R., Jones L.P., Barker J.S.F., 1968. The effects of population size and selection intensity in selection for a quantitative character in Drosophila: I. Short-term response to selection, Genet. Res. Camb., 12, 237-248.
Fuller J.L. 1969. The Genetics of behavior, in The Behavior of Domestic Animals, E.S.E. Hafez (ed.), Balliere, Tindall and Cassel ltd., London, pp. 45-64.
Hale E. В., 1969. Domestication and the evolution of behavior. In: The Behavior of Domestic Animals, E. S. E. Hafez (ed.), Baillierc, Tindall and Cassell Ltd., London, pp. 22-44.
Hammond K., 1973. Population Size, Selection Response and Variation in Quantitative Inheritance. Ph. D. thesis, Sydney University.
Hanrahan I.P., Eisen E.J., Legates I.E., 1973. Effects of population size and selection intensity on short-term response to selection for post weaning gain in mice, Genetics, 73, 513-530.
Harlan J.R., 1975. Our vanishing genetic resources, Science, 188, 618-621.
Kimura M.,1965. A stochastic model concerning the maintenance of genetic variability in quantitative characters, Proc. Natl. Acad. Sci. U.S.A., 54, 731-736.
Kimura M., Crow J.F.,1963. The measurement of effective population number. Evolution, 17, 279-288.
Lack D., 1954. The Natural Regulation of Animal Numbers, Clarendon Press, Oxford.
Lande R., 1976. The maintenance of genetic variability by mutation in a polygenic character with linked loci, Genet. Res. Camb.. 26, 221-235.
Latter B.D.H., 1970. Selection in finite populations with multiple alieles: II. Centripetal selection, mutation and isoallelic variation, Genetics, 66, 165-186.
Latter B.D.H., Robertson A., 1962. The effects of inbreeding and artificial
selection on reproductive fitness, Genet. Res. Camb., 3, 110-138.
Lerner I.M., 1954. Genetic Homeostasis, Oliver and Boyd Ltd., Edinburgh.
Lewontin R.C, 1978. Adaptation, Scientific American, 239, 156-169.
Nei M., Maruyama Т., Chakrabotry R., 1975. The bottleneck effect and genetic variability in populations, Evolution, 29, 1-10.
Prout Т., 1962. The effects of stabilizing selection on the time of development in Drosophila melanogaster, Genet. Res., 3, 364-382.
Rendel J.M., 1943. Variations in the weights of hatched and unhatched duck's eggs, Biomatrica, 33, 48-56.
Rendel J.M., 1960. Selection for canalization of the scute phenotype in Drosophila melanogaster, Aust. J. Biol. Sci., 13, 36-47.
Sved J.A., Ayala F.J.,1970. A population cage test for heterosis in Drosophila pseudoobscura, Genetics, 66, 97-113.
Waddington С.H.,1957. The Strategy of the Genes, Allen and Unwin, London.
Weldon W.F.R., 1901. A first study of natural selection in Clausilia laminata (montague), Biometrika, I, 109-124.
Wright S., 1977. Evolution and the Genetics of Populations, Vol. 3, Experimental Results and Evolutionary Deductions, University of Chicago Press, Chicago.