




Мухина Е.А. Содержание джека в Бухарском питомнике и некоторые особенности постэмбрионального развития его птенцов // Дичефермы и зоопитомники. Сборник научных трудов ЦНИЛ Главохоты РСФСР. М., 1991. С. 129-143.
В настоящее время для многих видов дрофиных (Otididae), которые находятся на грани исчезновения (стрепет, дрофы - большая, индийская, австралийская, некоторые виды американских) созданы специализированные питомники и центры по разведению в неволе.
Разработки по разведению в неволе джека (Chlamydotis undulata) ведутся в научно-исследовательских центрах Израиля (Mendelssohn, 1979, 1982), Саудовской Аравии (Launay, Paillat, 1990). В Советском Союзе этот вид содержался в некоторых зоопарках (Синявский, 1985). В Бухарском питомнике первые попытки его содержания были предприняты в 1980 г. (Флинт и др., 1982; Пономарева, 1983), планомерное же изучение развернулось в 1983 г., когда для содержания джека были построены вольеры площадью 34х38 м2 из металлической сетки с ячеей 100x100 мм. Оказалось, что это ограждение непригодно, так как через такую ячею в вольеры могли проникать мелкие хищники. В 1986 г. вольеры были обтянуты по периметру звероводческой сетью, диаметр ячеи которой был 10х30 мм и 15х15 мм. Позднее кроме устройства дополнительных секций по периметру выделили четыре внутренних секции, в центре которых установили юрту, используемую под зимовальное помещение, построили наблюдательный пункт и установили дезбарьер на входе. Все секции, в которых содержались птицы, были закрыты сеткой - сверху шелковой и хлопчатобумажной, а угловые - металлической. Чтобы хищники не проникали внутрь через верх, по периметру был установлен наружный козырек из проволоки с натянутой на нее шелковой сетью, которая на следующий год была заменена металлической. В настоящее время вольеры разделены на 12 секций, площадью от 42 до 175 кв. м. Высота сети ограждения 2 м, она прикреплена к бетонным столбикам. Во избежание подкопов сеть снизу забетонирована и закопана на глубину до 40 см. На верхней части установлен козырек из металлической сети с отрицательным наклоном наружу, его ширина 60-70 см. Снизу ограждение вольер закрыто тростниковыми щитами, ограничивающими видимость и создающими дополнительные укрытия. Схема вольер представлена на рис. 1.
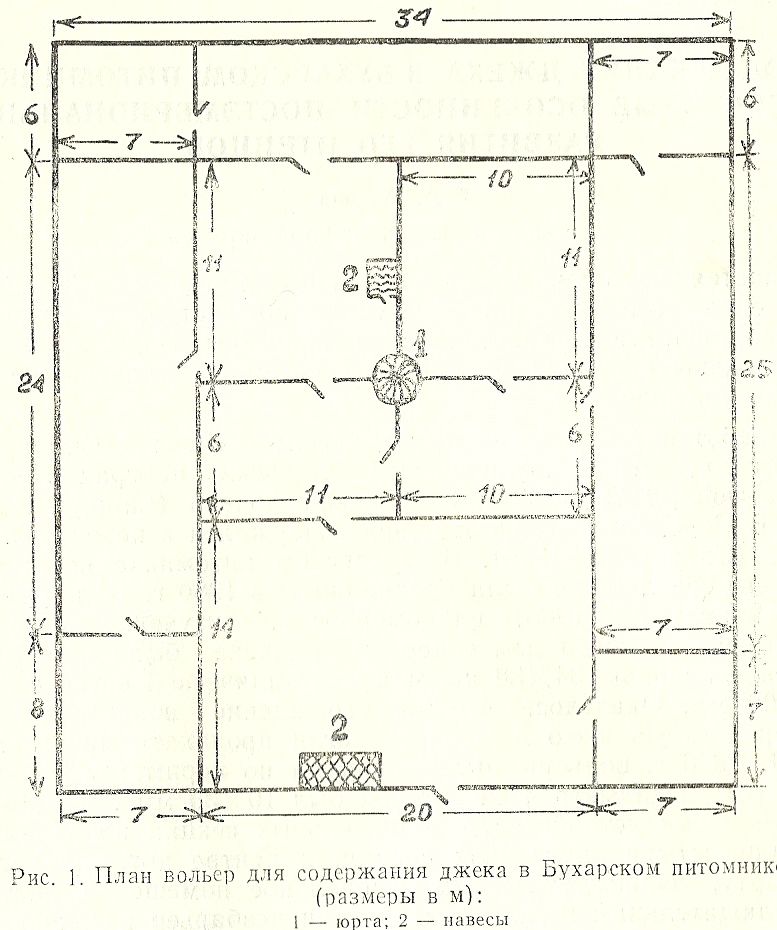
Рис. 1. План вольер для содержания джека в Бухарском питомнике (размеры в м): 1 - юрта; 2 – навесы.
Вольеры граничат со свободной территорией, откуда в питомник постоянно могут проникать хищники, потери от них происходили не один раз. В 1983 г. в начале июня в вольеру через крупную ячею проникла перевязка, задушила взрослого джека, а в конце июня - забралась к 3 птенцам и все они погибли. В 1988 г. в июле в вольеру через верх забралась степная кошка и задушила одного из молодых самцов, а в 1989 г. в октябре в вольеру также через верх попал этот же хищник, предварительно прорвав шелковую сеть козырька, после чего шелковый козырек был заменен металлическим. Дополнительно вдоль вольер расставляли капканы, в которые в осенне-зимний период 1989/90 г. попалось 6 степных кошек.
Это свидетельствует о том, что защита от хищников при разведении птиц всегда актуальна и при проектировании новых вольер в питомниках обязательно нужно предусматривать надежные перекрытия сверху.
Группы птенцов или взрослых птиц можно содержать вместе, между ними конфликты не возникают. Однако, взрослых особей и птенцов до 3-месячного возраста необходимо разделять, т. к. временами взрослые начинают преследовать птенцов и не подпускают их к корму.
Площадь вольер при содержании птенцов и взрослых птиц в нерепродуктивный период большого значения не имеет. В нашем питомнике птенцы и взрослые птицы по 7 особей подолгу содержались в одной небольшой секции, площадью 7x6 м2 и между ними не возникало территориальной напряженности. Но лучше все-таки содержать их в просторных вольерах, так как в этом случае полнее удовлетворяются физиологические и социальные потребности птиц. Кроме того, по-видимому, желательно разделять взрослых джеков на пары еще с осени (Mendelssohn, 1979), т. к. при содержании их в одной группе в течение года могут быть нарушены механизмы размножения. По опыту нашей работы, большая часть самок при этом вообще не приступает к гнездованию, либо же откладывает неоплодотворенные яйца, это же наблюдалось и в экспериментах Тель-Авивского научно-исследовательского центра.
В Бухарском питомнике вольеры построены на участке песчаной пустыни с присутствием нескольких кустов черного саксаула, естественную растительность при постройке не нарушили. Кроме того, вдоль стенок, разделяющих секции, были установлены тростниковые щиты, которые создали дополнительные укрытия от солнечных лучей и снизили фактор беспокойства. Птицы пользуются ими в основном в жаркое время года.
Для предотвращения травм, чаще всего получаемых при попытках взлета, у всех птиц (в том числе и у птенцов, начиная с 2-месячного возраста) подрезали маховые перья на одном из крыльев, повторяя операцию по крайней мере два раза в год.
При достаточном обеспечении необходимыми кормами, джеки неплохо зимуют в питомнике, так как зима под Бухарой обычно довольно сухая, с небольшим количеством действительно холодных дней.
Они легко переносят резкое снижение температуры, если оно не сопровождается выпадением атмосферных осадков, при которых защитные свойства оперения джеков резко снижаются и возможно переохлаждение птиц. При продолжительном снегопаде, особенно если он с дождем, оперение джеков намокает, они слабеют, начинают садиться или ложиться в снег. В такие периоды использовали юрту, в которую было подведено электричество и внутри устроен подогреваемый пол. Самостоятельно джеки в юрту не заходили, приходилось каждый раз их отлавливать что сопряжено со стрессами для птиц. В 1989 г. нами был использован более щадящий способ загона: два загонщика с куском натянутой между ними сети, постепенно подводили джеков к входу в юрту, сужая воронку.
Зимой 1989/90 г. джеки провели в юрте в общей сложности 25 дней. Их туда помещали только на ночь при сильных морозах (до -17-20 °С) или на несколько суток при снегопаде, снеге с дождем или дожде с последующим похолоданием. В юрте джеки располагались на подогреваемом полу. Мы заметили, что при похолоданиях, сопровождаемых выпадением осадков, у джеков часто были пищеварительные расстройства.
Для предотвращения инфекционных заболеваний и гельминтозов был установлен дезбарьер на входе, который один раз в месяц заправляется раствором едкого натра, а в этот промежуток поддерживается влажность; ограничен вход внутрь вольер; раз в неделю в вольерах проводили уборку накопившихся экскрементов, выпавших перьев, мусора, мыли кормушки; два раза в год производили дегельминтизацию птиц.
Кормление
Режим кормления для взрослых птиц в течение всего года был двухразовый - утренний и вечерний - совпадающий с ритмом их дневной активности. Птенцов в первую неделю жизни кормили через каждые два часа, затем до месячного возраста - 5, 4 и 3 раза в день, позднее переводили на двухразовый режим кормления.
При составлении рационов исходили из практического опыта и данных по питанию птиц в пустыне. Использовали комбикорм ПК-2 (основа рациона в течение всего года), мясо или рыбу (также в течение всего года при суточной норме не больше 30-40 г на 1 птицу), творог - 40 г в день на 1 особь, яйцо (вареное или сырое) в качестве добавки в составляемую смесь, зелень (люцерна, кукуруза, лебеда, парнолистник и др.), овощи (морковь, лук, помидоры, огурцы, болгарский перец, свекла, капуста и т. д.), сухие дрожжи - 0,5 г, поливитамины.
Кроме того, использовали «нестабильные корма»: насекомых, отловленных в период их ночного лета, а также ящериц. В вольерах джеки весной поедали всходы и проростки растений, почки и распускающиеся листья, цветы и свежие семена; осенью - семена солянок, чогона и саксаула. Отмечено, что разнообразный по компонентам корм поедается лучше и при этом не наблюдается отклонений в процессах пищеварения. В течение всего года джекам предлагали известняк, ракушечник и яичную, скорлупу, которые особенно хорошо поедались в периоды линьки.
Как показывает наш опыт, при указанных выше условиях невозможно длительное содержание джека в неволе. Но для разведения его в питомнике необходимы дополнительные меры, которые бы способствовали формированию нормальных социальных отношений в группе, необходимых для размножения. В вольерах питомника наблюдалось развитие репродуктивного поведения джека. В 1989-1990 гг. впервые в СССР отмечалось гнездование одной из самок джека в неволе. Она отложила в общей сложности 7 яиц: два в первый год и пять во второй. Однако, все яйца оказались неоплодотворенными. В 1990 г. самка села их насиживать, но яйца в гнезде подменили привезенными с сопредельной территории, что позволило пронаблюдать за вылуплением птенцов, их ростом и развитием.
Работ, посвященных постэмбриональному развитию птенцов джека, немного. В основном, это небольшие упоминания в обзорных статьях и общие характеристики (Mendelssohn, et al. 1979, 1982; Mendelssohn, 1982). В отечественной литературе общий анализ постэмбрионального развития птенцов джека представлен работой Т.С. Пономаревой (1983).
Нами предпринята попытка обобщить данные по росту и развитию птенцов джека в условиях вольер, как при искусственном выкармливании, так и в выводке. Рассматриваются процессы роста и развития разного пола, возможные отклонения в развитии, связанные с действием внешних факторов.
Материал собран при наблюдениях за 25 птенцами джека, выращенных в вольерах Бухарского питомника с 1983 до 1990 год. В 1983 г. наблюдения производились за 9 птенцами, 5 из которых вывелись при инкубировании яиц под курицей-наседкой, а 4 - отловлены на воле в возрасте 9-11 дней. Фиксировали размеры по 5 основным параметрам (масса тела, величина клюва, цевки, крыла и рулевых) до 55-65-дневного возраста. В 1986 г. наблюдения проводили за 4 птенцами, один из которых пойман однодневным, два - в возрасте около 20 дней, а еще один привезен в возрасте около 1,5 месяцев, сильно ослабленным в результате дистрофии. Измерения вели по 27 параметрам по методике, предложенной В.С. Ивановой (1982, 1986), с тем, чтобы проследить взаимосвязь в развитии разных частей тела. В 1986 г. все промеры регулярно снимали до 90-дневного возраста, сначала с периодичностью 3-5 дней (масса - ежедневно до 40 дней), затем - один раз в 7 дней (масса - 2 раза в неделю). Осенью дополнительно взвешивали птиц - один раз в месяц. В 1987 г. наблюдения проводили за 4 птенцами, два из которых отловлены в возрасте двух дней, один привезен в возрасте 40-50 дней сильно ослабленным и один - в возрасте 10-14 дней. Промеры производили один раз в 5-7 дней до 70-дневного возраста. В 1988 г. наблюдения вели за 3 птенцами, отловленными в возрасте двух дней. Первые 25 дней птенцов ежедневно взвешивали, а дальше - до 40 дней - через день и до 60 дней - один раз в 5 дней. Промеры по остальным параметрам производили один раз в 5-6 дней. В 1990 г. наблюдения вели за 5 птенцами, 3 из которых выведены из яиц, инкубировавшихся под насиживающей самкой, позднее выращивающей их, один - в инкубаторе и один привезен в возрасте 6-8 дней. До 14 дней промеры были сделаны 4 раза, затем один раз в 7 дней до возраста 85 дней у птенцов из выводка.
Все птенцы были помечены индивидуальными цветными пластиковыми кольцами. Взвешивание проводили на технических весах с точностью до 0,1 г. По мере подрастания птенцов и увеличения их беспокойства при измерительных процедурах точность на весах выдерживалась до 1 г. Остальные параметры снимали при помощи штангенциркуля, мягкой мерной ленты и линейки.
Птенцы джека появляются на свет зрячими (радужина глаз светлая, цвет ее несколько варьирует от светло-серого до светло-желтого), с открытыми слуховыми проходами, тело покрыто густым беловато-желтым пухом, с темными пятнами, имеющими определенный рисунок. На каждом предплечье имеется желтое пятно в черной окантовке, на голове с каждой стороны по три симметричных пятна, на спине хорошо выражены четыре крупных пятна и несколько мелких. Все пятна в центре желтовато-коричневые, а по краям с темной, почти черной окантовкой. Низ тела - грязно-белый. Хорошо выражена шейная аптерия длиной до 30 мм. Плюсны голубовато-серые, а ступни ног желтовато-розовые. По мере увеличения массы тела птенцов, пух на них бледнеет, по-видимому, за счет того, что пушинки расходятся и частично разворачиваются. При неравномерном росте птенцов в одном выводке разница между ними в первые дни резко заметна - птенцы, отстающие в росте, намного ярче, чем опережающие их собратья.
Первостепенные маховые появляются на 4-6 день; чаще всего на 5 день (8 птенцов), на 4-й - (один в 1986 г.) и на 6-й - (один в 1990 г.). Пеньки маховых появляются на месте пушинок и уже несут сверху кисточку-пушинку, которая сначала не растет или растет очень слабо. Пеньки первых маховых растут до 20-25 дней, достигают размеров 30-45 мм, а затем начинают крошиться. Бурный рост кисточек (опахал) начинается на 14-15 день и продолжается до 28-30 дней, после чего происходит некоторое замедление, однако, маховые продолжают расти до 80-85 дней (начало линьки). Пеньки кроющих первых маховых появляются одновременно с маховыми, но кисточки на них начинают раскрываться на 8-11 день. Рост пеньков кроющих маховых продолжается до 14-22 дней, а опахал - до 28-35 дней. К этому времени кроющие маховых полностью сформировываются (табл. 1, 2). При сравнении роста птенцов в выводке и при искусственном выкармливании видно, что заметные расхождения в развитии оперения есть в первые 20 дней, затем они становятся незначительными. Рост маховых, как и остальных частей оперения, заметно варьирует на ранних стадиях онтогенеза в зависимости от индивидуальных особенностей птенцов, также как и от их состояния.
Таблица 1
Рост различных частей тела и оперения у птенцов джека при искусственном выкармливании (средние значения)
|
Показатели |
Возраст (дни) |
|||||||
|
7 |
10 |
14 |
16 |
20 |
23 |
25 |
28 |
|
|
Длина голени |
42 |
45 |
56 |
63 |
65 |
71 |
- |
75 |
|
Длина бедра |
34 |
35 |
41 |
45 |
45 |
51 |
- |
56 |
|
Длина когтя |
3,5 |
3,5 |
3,5 |
- |
4,2 |
4,6 |
- |
4,6 |
|
Длина среднего пальца |
15 |
15,5 |
20 |
22 |
24 |
27 |
- |
27 |
|
Длина предплечья |
40 |
43 |
52 |
65 |
71 |
82 |
- |
98 |
|
Длина плеча |
31 |
39 |
46 |
55 |
57 |
68 |
- |
82 |
|
Длина кисти |
31 |
31,6 |
40 |
50 |
56 |
71 |
- |
78 |
|
Длина головы |
29 |
33 |
36 |
36 |
42 |
42 |
- |
45 |
|
Ширина головы |
23 |
25 |
28 |
28 |
28 |
30 |
- |
32 |
|
Высота головы |
22 |
26 |
26,5 |
26,5 |
27,6 |
30 |
- |
31 |
|
Длина головы с клювом |
50 |
52 |
57 |
61 |
63 |
69 |
- |
74 |
|
Кроющие рулевых: |
|
|
|
|
|
|
|
|
|
пенек |
1,7 |
1,7 |
4,7 |
9 |
10 |
- |
10,3 |
10,5 |
|
опахало |
0 |
0 |
3,3 |
11 |
19 |
- |
34,7 |
42 |
|
Крайние рулевые: |
|
|
|
|
|
|
|
|
|
пенек |
0 |
1 |
7 |
11 |
17 |
- |
19 |
19,5 |
|
опахало |
0 |
0 |
3 |
13 |
22 |
- |
33 |
46 |
|
Средние рулевые: |
|
|
|
|
|
|
|
|
|
пенек |
1,8 |
3 |
7 |
14 |
20 |
- |
25 |
34 |
|
опахало |
0 |
0 |
3,7 |
9 |
18 |
- |
35 |
36 |
|
Кроющие 1-х маховых: |
|
|
|
|
|
|
|
|
|
пенек |
11 |
11 |
20 |
21 |
22 |
- |
18 |
16 |
|
опахало |
0,5 |
0,5 |
11 |
22 |
46 |
- |
72 |
72 |
|
1-ые маховые: |
|
|
|
|
|
|
|
|
|
пенек |
18 |
21 |
31 |
34 |
36 |
- |
39 |
37 |
|
опахало |
1 |
4 |
9 |
25 |
47 |
- |
66 |
94 |
Продолжение таблицы 1
|
Показатели |
Возраст (дни) |
||||||||
|
30 |
32 |
36 |
40 |
45 |
50 |
60 |
70 |
85 |
|
|
Длина голени |
- |
84 |
88 |
98 |
109 |
110 |
118 |
- |
128 |
|
Длина бедра |
- |
67 |
65 |
68 |
71 |
74 |
74 |
- |
86 |
|
Длина когтя |
- |
5,1 |
5,4 |
6 |
6,6 |
6,6 |
8 |
- |
8,7 |
|
Длина среднего пальца |
- |
29 |
29,5 |
31 |
32 |
32 |
34 |
- |
36 |
|
Длина предплечья |
- |
105 |
111 |
121 |
134 |
135 |
139 |
- |
144 |
|
Длина плеча |
- |
89 |
95 |
103 |
112 |
114 |
117 |
- |
119 |
|
Длина кисти |
- |
91 |
104 |
111 |
116 |
118 |
123 |
- |
128 |
|
Длина головы |
- |
48 |
50 |
50 |
- |
54 |
59 |
- |
61 |
|
Ширина головы |
- |
33 |
34 |
36 |
- |
37 |
38 |
- |
38 |
|
Высота головы |
- |
32 |
32 |
34 |
- |
36 |
36 |
- |
36 |
|
Длина головы с клювом |
- |
76 |
79 |
82 |
- |
91 |
93 |
- |
100 |
|
Кроющие рулевых: |
|
|
|
|
|
|
|
|
|
|
пенек |
10,8 |
- |
6,8 |
7,5 |
- |
5 |
- |
- |
- |
|
опахало |
51 |
- |
57 |
66 |
- |
71 |
- |
- |
- |
|
Крайние рулевые: |
|
|
|
|
|
|
|
|
|
|
пенек |
23 |
- |
23 |
24 |
19,5 |
19 |
14 |
4 |
- |
|
опахало |
54 |
- |
63 |
85 |
97 |
119 |
139 |
141 |
164 |
|
Средние рулевые: |
|
|
|
|
|
|
|
|
|
|
пенек |
31 |
- |
33 |
26 |
22 |
18 |
16 |
10 |
- |
|
опахало |
52 |
- |
67 |
94 |
114 |
126 |
150 |
- |
171 |
|
Кроющие 1-х маховых: |
|
|
|
|
|
|
|
|
|
|
пенек |
14 |
- |
11 |
11 |
11 |
6 |
4 |
- |
- |
|
опахало |
73 |
- |
74 |
75 |
75 |
75 |
76 |
- |
76 |
|
1-ые маховые: |
|
|
|
|
|
|
|
|
|
|
пенек |
33 |
- |
31 |
24 |
19 |
18 |
16 |
12 |
9 |
|
опахало |
103 |
- |
122 |
151 |
181 |
193 |
219 |
231 |
239 |
Таблица 2
Рост различных частей тела и оперения у птенцов джека в выводке
|
Показатели |
Возраст (дни) |
||||||||||
|
6 |
8 |
11 |
14 |
21 |
28 |
36 |
43 |
50 |
57 |
71 |
|
|
Длина голени |
36 |
40 |
45 |
52 |
64 |
77 |
85 |
99 |
- |
107 |
117 |
|
Длина бедра |
26 |
32 |
35 |
39 |
49 |
54 |
59 |
65 |
- |
67 |
68 |
|
Длина когтя |
3,4 |
3,5 |
4 |
4 |
5 |
5 |
6 |
7 |
- |
8 |
8 |
|
Длина среднего пальца |
14 |
15 |
16 |
20 |
23 |
26 |
29 |
31 |
- |
36 |
36 |
|
Длина предплечья |
29 |
30 |
37 |
47 |
68 |
90 |
101 |
115 |
- |
132 |
136 |
|
Длина плеча |
25 |
26 |
34 |
44 |
60 |
80 |
90 |
104 |
- |
109 |
122 |
|
Длина кисти |
26 |
30 |
34 |
45 |
70 |
90 |
102 |
121 |
- |
127 |
145 |
|
Длина головы |
28 |
29 |
32 |
35 |
38 |
41 |
48 |
54 |
- |
59 |
65 |
|
Ширина головы |
24 |
26 |
26 |
29 |
31 |
31 |
33 |
35 |
- |
38 |
40 |
|
Высота головы |
21 |
22 |
23 |
25 |
28 |
30 |
32 |
33 |
- |
35 |
37 |
|
Длина головы с клювом |
46 |
51 |
53 |
58 |
65 |
72 |
79 |
88 |
- |
95 |
103 |
|
Кроющие рулевых: |
|
|
|
|
|
|
|
|
|
|
|
|
пенек |
- |
- |
- |
- |
- |
24 |
8 |
- |
- |
- |
- |
|
опахало |
- |
- |
- |
- |
- |
46 |
50 |
66 |
- |
- |
70 |
|
Крайние рулевые: |
|
|
|
|
|
|
|
|
|
|
|
|
пенек |
- |
- |
- |
- |
7 |
11 |
27 |
24 |
23 |
20 |
- |
|
опахало |
- |
- |
- |
- |
18 |
43 |
64 |
98 |
122 |
144 |
175 |
|
Средние рулевые: |
|
|
|
|
|
|
|
|
|
|
|
|
пенек |
- |
- |
3 |
9 |
17 |
25 |
27 |
24 |
21 |
21 |
- |
|
опахало |
- |
- |
3 |
4 |
24 |
49 |
70 |
105 |
130 |
142 |
175 |
|
Кроющие 1-х маховых: |
|
|
|
|
|
|
|
|
|
|
|
|
пенек |
- |
8 |
16 |
21 |
17 |
10 |
10 |
9 |
- |
- |
8 |
|
опахало |
- |
3 |
4 |
10 |
48 |
68 |
70 |
71 |
- |
- |
73 |
|
1-ые маховые: |
|
|
|
|
|
|
|
|
|
|
|
|
пенек |
4 |
10 |
17 |
27 |
35 |
35 |
22 |
20 |
- |
15 |
- |
|
опахало |
3 |
4 |
6 |
13 |
50 |
89 |
136 |
177 |
- |
213 |
236 |
Рулевые начинают расти на 10-14 день; при нормальном развитии птенца пеньки рулевых появляются уже на 10-й день, а при задержках, связанных с переохлаждением и заболеваниями - несколько позднее (в 1988 г. у птенца, перенесшего диспепсию в 8-11-дневном возрасте, рулевые появились на 14-й день). Сначала быстро растут пеньки, опахала же медленно, их рост начинается после 20-25 дней, полностью они раскрываются на 45-50 день, хотя рост рулевых, как и рост маховых, продолжается до начала линьки.
Кроющее оперение появляется сначала на крыле. На 7-8 день жизни птенцов образуются пеньки кроющих на крыльях, по сторонам от лопаток. Перо растет из отдельных пушинок, которые остаются на вершинах пеньков, а затем разворачивающихся опахал небольшими слегка раскрывающимися кисточками. Пух, который не переходит в перо, также претерпевает изменения; пушинки делятся на мелкие бороздки, разворачиваются «кустиком» и бледнеют. Через 1-2 дня после появления пеньков кроющих на крыле начинают расти пеньки кроющих на спине - сначала вверху, затем посередине и внизу (по будущим птерилиям). Одновременно у птенцов вытягиваются шея и ноги. На голове пух сохраняется примерно до 20 дней, а пеньки контурных перьев появляются на 20-22 день: сначала над бровями - двумя линиями, затем по центру. После того, как начинает расти контурное оперение на голове, становится заметна замена пуха и по бокам шеи (25-30 день). Там образование контурного пера также осуществляется поэтапно: сначала двумя полосами по бокам, затем - впереди. К месячному возрасту формирование контурного оперения в основном заканчивается, пух сохраняется отдельными полосками по бокам тела, головы, шеи и подмышками. Шейная аптерия на затылке зарастает коротким пухом, едва выраженным в межлопаточной области. Перо уже плотно уложено на крыле и на спине, основное поле пера - песочно-охристое. По всему перу четко выражена тонкая струйчатость. Хвост, как и у взрослых, более охристый, чем оперение спины. Тонкая светлая полоска на кроющих крыла совсем белая, на маховых — слегка охристая с редкими черными точками. Пух сохраняется на видимых частях тела отдельными тонкими полосками до 40-50 дней, под крылом и по бокам тела он держится у отдельных птенцов до 80-85 дней. У птенцов, в отличие от взрослых, контурное кроющее ювенильное оперение спины более светлое за счет светлых каплевидных пятен, которыми заканчиваются кроющие верха тела. К 1,5 месяцам формирование оперения у птенцов в основное заканчивается и тут же отмечается выпадение отдельных кроющих перьев - начинается линька. К 1,5 месяцам птенцы приобретают пропорции взрослых птиц, но рост отдельных частей тела еще продолжается до осени.
Половой диморфизм начинает проявляться уже в возрасте 30-35 дней, что отмечено в некоторых работах (Mendelssohn, et. аl., 1982), но расходится с наблюдениями Т.С. Пономаревой (1983). Выявить среди птенцов самцов и самок вначале довольно сложно, их различие проявляется в неравномерном росте частей тела. Их можно различать по появлению перьев воротничка - у самцов они заметны раньше, чем у самок - на 33-35 день, и сразу растут более широкой темной полоской. В 1990 г. по этому признаку мы верно определили пол птенцов на 35-й день. Кроме того, у птенцов-самцов в возрасте 45-50 дней на голове образуется светлая шапочка из перьев с белыми основаниями и пестрыми вершинками, у самок такой шапочки нет, она появляется у них обычно только на второй год жизни. Зато у самочек к 1,5-месячному возрасту в нижней части шеи развивается ошейник, состоящий из расширенных серовато-голубоватых перьев. Различие между самцами и самками быстро увеличивается и в два месяца становится очевидным, так как самцы продолжают расти несколько быстрее и резко обгоняют самок в размерах. Двухмесячные самцы ростом с водящую выводок самку, а в 80 дней они уже выше ее за счет длинных ног и шеи; самочки же лишь к 80-85 дням достигают размеров взрослой самки, но долго еще кажутся меньше ее вследствие меньшей массы тела. В возрасте около 80 дней у самцов еще начинают формироваться белые и черно-белые перья воротника, которые у молодых самок вообще не выражены. Линька начинается в возрасте около 2-х месяцев, на 3-4-й месяцы сменяется контурное оперение крыла, верха и низа тела. В 80-85 дней начинают сменятся маховые перья, последовательно от внутренних к наружным: сначала первостепенные 7, 8, 9 и их кроющие, линька идет синхронно у одновозрастных птенцов с небольшими индивидуальными различиями, одновременно на обоих крыльях.
Нарастание массы тела идет непрерывно со 2-4-го дня жизни, хотя возможны и отдельные задержки в начале роста (слабые птенцы в 1983, 1987, 1988 и 1990 гг.), когда из-за болезни или переохлаждения наблюдается замедление роста и даже понижение массы тела. Если в возрасте до 30 дней различия в массе тела птенцов обоих полов связаны только с особенностями индивидуального развития, определяемыми внешними условиями, то в месячном возрасте и старше максимальные величины характерны только для самцов, а минимальные - для самок. Резкое расхождение в увеличении массы тела начинает проявляться у самцов и самок в возрасте около месяца и дальше это различие постепенно увеличивается, так что к двум месяцам масса тела самца заметно больше массы тела самки. Расхождения в росте различных частей тела (опережение у самцов) становятся заметны позднее - к 40-50-дневному возрасту.
При выкармливании птенцов человеком увеличение массы тела происходит неравномерно, скачкообразно, но стабильно, а рост отдельных частей тела резко идет вначале (до 30-40 дней), а затем замедляется. В выводке рост птенцов протекает более «спокойно», без резких отклонений и скачков. Однако в целом средние размеры птенцов в выводке меньше, чем у одновозрастных птенцов, выращиваемых человеком. Вероятно, такое различие определялось соотношением полов (в выводке оно было 1:3 - 1 самец и 3 самки, а для остальной выборки - приблизительно - 2:1). Но возможны и другие причины, например, состав кормов и активность птенцов. Птенцы, выкармливаемые человеком, были менее подвижны, к тому же им предлагался более калорийный корм (насекомые, мясо, яйца, рыба), птенцы же в выводке были более активными, так как много перемещались вслед за самкой, которая предлагала им корм, бедный протеиновыми компонентами и более богатый растительными.
Скорость роста (Шмальгаузен, 1935) носила скачкообразный характер, наблюдались индивидуальные различия, но прослеживаются и общие закономерности. В первые дни после вылупления скорость роста птенцов низка (это период «адаптации» к внешней среде, когда еще не рассосался желточный мешок), на 3-5 день начинается бурный рост массы тела, который протекает неравномерно, но характеризуется одним или двумя резкими пиками до 20-дневного возраста, после чего заметен резкий спад и колебание в пределах одного уровня. После 30-дневного возраста становится заметной стабильно большая скорость роста массы тела у самцов. Быстрый рост массы тела коррелирует с высокой скоростью роста крыла и цевки, для которых наибольшие величины характерны в течение первых 20-25 дней, а затем заметна тенденция к снижению, которое происходит постепенно и довольно плавно с 30 до 50-55-дневного возраста, после чего скорость роста этих частей тела становится очень низкой и приближается к нулю.
Скорость роста первых маховых максимальна в первые дни после их появления, затем заметен довольно резкий спад, а между 14-16 днем развития - снова резкий скачок вверх, после которого наблюдается, уже устойчивое снижение скорости роста и ее незначительные колебания на одном уровне. Она замедляется, но не прекращается до 65-75-дневного возраста птенцов, т. е. рост маховых продолжается до самой линьки. Рулевые начинают расти через 5-7 дней после первых маховых и скорость их роста также, в первые дни, протекает бурно, а затем постепенно и плавно снижается. Максимальна скорость роста рулевых на 11-14 день. Сначала до 20 дня идет резкий спад, затем постепенное плавное снижение до 25-28 дня и затем - колебания на очень низком уровне. Рост рулевых, также, как и маховых, прекращается перед самой линькой.
Наши данные об удельной скорости роста отличаются от тех, которые представлены в работе Т.С. Пономаревой (1983), и различаются для разных групп и отдельных птенцов, что свидетельствует о большой зависимости скорости роста от индивидуальных особенностей птенцов, их физиологического состояния, а также от действия внешних факторов (питание, активность, болезни). Кроме того, она зависит от пола птенцов. По нашим наблюдениям, удельная скорость роста самца и самки из выводка до 30-дневного возраста характеризуется общими закономерностями и существующие различия определяются индивидуальными: особенностями роста птенцов (так, рост рулевых у самца начался позднее - на 12 день, но протекал более бурно, чем у самки). Зато с возраста 30-35 дней заметна явная тенденция к большей скорости роста у самца, которая прослеживается по различным параметрам и устойчиво держится до замедления общей скорости роста к 60-80 дням.
Рост оперения прекращается с началом линьки, поэтому скорость роста рулевых и первых маховых замедляется и постепенно падает до нуля с 60-дневного возраста до 85 дней, медленный рост остальных частей тела продолжается до осени, но при очень низкой (близкой к нулю) скорости. Нарастание массы тела происходит вплоть до октября, т. е. почти до 6 месяцев. Самцы к этому времени достигают массы тела 1,6-1,9 кг, самки - 0,9-1,2 кг.
Константа роста вычислялась нами по формуле Шмальгаузена (1935). Увеличение удельной скорости роста и соответственно константы роста идет с 5 до 10 дней (табл. 3). Наибольшие скорость и константа роста отмечены с 11 по 15 день. С 16 по 40 день идет постепенное снижение скорости роста, а для константы роста в этот период характерны небольшие колебания на одном уровне - от 1,026 до 1,265. Дальнейший переход (с 41 до 85 дней) характеризуется постепенным снижением скорости роста до нуля и резкими неравномерными колебаниями константы роста.
Таблица 3
Удельная скорость (С) и константа роста (К) на разных стадиях постэмбриогенеза у птенцов джека (n=20)
|
Возраст, дни |
С |
К |
Возраст, дни |
С |
К |
|
1-5 |
0,063 |
0,189 |
41-45 |
0,023 |
0,98 |
|
6-10 |
0,083 |
0,623 |
46-50 |
0,028 |
1,33 |
|
11-15 |
0,132 |
1,716 |
51-55 |
0,023 |
1,21 |
|
16-20 |
0,057 |
1,026 |
56-60 |
0,041 |
2,36 |
|
21-25 |
0,046 |
1,058 |
61-65 |
- 0,014 |
- 0,875 |
|
26-30 |
0,046 |
1,265 |
66-70 |
0 |
0 |
|
31-35 |
0,035 |
1,155 |
71-75 |
0,018 |
1,305 |
|
36-40 |
0,029 |
1,102 |
76-85 |
0,005 |
0,4 |
Мы сопоставили величины скорости и константы роста в разных группах птенцов - по средним значениям для птенцов из выводка и выкармливаемых человеком, а также по максимальным и минимальным параметрам для возраста от 1 до 70 и от 1 до 85 дней (табл. 4). Интересно, что удельная скорость роста птенцов в выводке и выращиваемых человеком совпадает, хотя процесс роста птенцов первой группы более замедленный и до 70 дней их скорость роста ниже, чем у птенцов второй группы. Кроме того, совпадает скорость роста самцов с максимальной скоростью роста, а самок - с минимальной в период с 1 по 85 день. И, соответственно, скорость роста самцов выше средней скорости роста и скорости роста самок. Скорость же роста самок ниже средней скорости роста; константы роста самцов имеют максимальные значения, а самок - минимальные.
Таблица 4
Соотношение удельной скорости роста и константы роста у разных групп птенцов джека за период с 1 по 70 и с 1 по 85 дни развития
|
Возраст, дни |
Птенцы, выкормленные человеком |
Птенцы из выводка |
||||
|
С ср. |
С макс. |
С мин. |
С ср. |
С самцов |
С самок |
|
|
1-70 |
0,044 |
0,045 |
0,043 |
0,041 |
0,043 |
0,038 |
|
К/1-70 |
1,562 |
1,60 |
1,53 |
1,48 |
1,57 |
1,37 |
|
1-85 |
0,036 |
0,038 |
0,036 |
0,036 |
0,038 |
0,034 |
|
К/1-85 |
1,548 |
1,634 |
1,56 |
1,53 |
1,61 |
1,46 |
Таким образом, удельная скорость роста птенцов джека мала только в первые три дня жизни, да еще в тех случаях, когда возможны какие-то отклонения в развитии, обусловливающие задержки роста. Дальше же - с 4 до 15-17 дней — идет интенсивный бурный рост, характеризующийся резким увеличением массы тела, ростом различных частей и стремительным формированием контурного оперения. В этот период удельная скорость роста различных частей тела держится на высоком уровне. Увеличение интенсивности скорости роста у птенцов джека начинается раньше, чем у других выводковых птиц - на 4-5-й день жизни, что может быть связано с адаптацией этого вида к обитанию в аридных условиях. По величине константы, возможно, довольно трудно выделить определенные закономерности в развитии и определить отдельные этапы роста птенцов, однако по величине удельной скорости роста они прослеживаются довольно четко. Можно выделить четыре этапа роста птенцов джека.
Период акклиматизации (Артамонова, 1969) — от вылупления до 3-5-дневного возраста. Характеризуется неустойчивостью птенцов к внешним воздействиям, низкой интенсивностью процессов роста. В первые два дня у отдельных птенцов происходит даже некоторое понижение веса. За этот период птенец адаптируется к внешним воздействиям и начинается его рост.
Период интенсивного роста и развития. По нашим данным, происходит с 5 до 20-25-дневного возраста. До 17-20 дней идет бурное нарастание массы тела, с 5 до 20 дней происходит интенсивный рост крыла и первостепенных маховых, а также остальных частей тела, с 10 до 25 дней - интенсивный рост рулевых перьев.
Период постепенного спада скорости роста. Длится с 25 до 40 дней и характеризуется постепенным замедлением роста разных частей тела. У самцов и самок этот процесс протекает по-разному, у самок скорость роста замедляется быстрее и держится затем на более низком уровне, чем у самцов.
Период равномерного медленного роста. Продолжается с 40 до 65-85 дней (а для массы тела - до осени), характеризуется незначительными колебаниями скорости роста па нижних пределах (кривые скорости роста разных частей тела на этом этапе сближаются).
Наблюдения за ростом и развитием разных групп птенцов при выкармливании их человеком и в выводке показывают, что, несмотря на некоторое расхождение в темпах роста на отдельных этапах, в целом, процесс роста от момента появления на свет до приобретения самостоятельности (85 дней) протекает сходно и характеризуется одинаковыми величинами удельной скорости роста и близкими константами роста для птенцов одного пола. Большое значение имеет сочетание эндогенных факторов, генетически обуславливающих процессы роста, и экзогенных - воздействия внешней среды. В частности, характер и интенсивность питания, которые приводят к резкому скачкообразному росту птенцов, выкармливаемых искусственно, в то время как в естественных условиях этот рост происходит более плавно и постепенно.
Адаптация к условиям аридного климата в ходе постэмбриогенеза проявляется в быстром развитии птенцов - высокой удельной скорости роста массы тела и различных его частей, стремительном формировании контурного оперения, быстрой самостоятельности, приобретении различных морфологических приспособлений, например, формирование на подошве джеков многослойного эпидермиса, который разрастается сосочковыми слоями и делает ступню нечувствительной к воздействию напочвенных температур, что позволяет легко перемещаться по раскаленной почве.
Установление пола молодых. Джеков возможно уже в возрасте 30-35 дней; более точное определение можно произвести в возрасте 45-50 дней, когда на голове у молодых самцов начинает образовываться шапочка из светлых перьев, не выраженная у молодых самок. Таким образом, половой диморфизм у джека проявляется относительно рано. По мере дальнейшего роста птенцов морфологические различия между самцами и самками постепенно возрастают и в 80-85 дней уже хорошо проявляются: это различие в массе тела, в размерах и пропорциях отдельных его частей, в наличии у самцов украшающих перьев воротника (черных и белых, тогда как у самок — только черных) и выделяющихся перьев шапочки. Особенных различий в поведении самцов и самок в этот период не наблюдается.
Таким образом, высокая скорость роста птенцов, формирующиеся у них морфологические приспособления и раннее созревание (в том числе и раннее распределение полов) во многом обеспечивают репродуктивный успех джека в экстремальных условиях аридного климата южных пустынь.
ЛИТЕРАТУРА
Артамонова З.В. Некоторые особенности постэмбрианального развития птиц в юго-восточных Каракумах // Зоол. журн. 1969. Т. 48. Вып. 11, С. 1706-1715.
Губин Б.М. Скляренко С.Л. Токовое поведение джека в восточных Кызылкумах // Известия АН КазССР. Серия биол. 1989. № 4. (154). С. 44-49.
Иванова В.С. Изучение развития молодняка некоторых видов дичи при искусственном разведении // Разведение и создание новых популяций редких и ценных видов животных: Тезисы докл. III совещ. Ашхабад, 1982. С. 66-73.
Иванова В.С, Трошкина Н.Н. Методические рекомендации по контролю за развитием молодняка птиц, разводимых в искусственных условиях. М., ЦНИЛ Главохоты РСФСР, 1986. 10 с.
Мухина Е.А. Бюджет времени и поведение джека в условиях вольер // Зоопарки и их роль в сохранении диких животных. Алма-Ата: Кайнар., 1989. С. 91-99.
Пономарева Т.С. Репродуктивное поведение и распределение джека в местах гнездования // Зоол. журн. 1983. Т. 62. Вып. 4. С. 592—601.
Пономарева Т.С. Постэмбриональное развитие птенцов джека // Зоол. журн. 1983. Т. 62. Вып. 8. С. 1221-1231.
Синявский А.В. О брачных играх джека // Сохраним диких животных. Алма-Ата, 1985. С. 117-118.
Тихонов Л.В. Акустическая сигнализация и поведение выводковых птиц (куриные и пластинчатоклювые) в раннем онтогенезе. 2. Сигнализация и поведение птенцов // Вестник МГУ. 1980. № 2. С. 3-12.
Флинт В., Пономарева Т., Прилипко Г. Джеки в вольере // Охота и охотн. хоз-во. 1982. № 5. С. 14-15.
Шмальгаузен И.И. Определение основных понятий и методика исследования роста // Рост животных. М.-Л.: Изд.-во АН СССР, 1935. С. 3-72.
Collar N.J. The Bustards and their conservation / 1982. Bustard in decline, Jaipur, 244-257.
Launay F., Paillat P. A behavioural repertoire of the adult Houbara bustard / Rev. Ecol. (Terre Vie), 1990. Vol. 45, p. 65-89.
Mendelssohn H. Development of Houbara population in Israel and Captive Breeding // The Houbara bustard. Athens, Greece. May 24. 1979.
Mendelssohn H., Marder U., Stavv M. Captive breeding of the Houbara and a description of its display // The International Council for Bird Preservation. XIII Bull. 1979. 134-149.
Mendelssohn H. Observations on the Houbara in Israel // Bustard in Decline. 1982. 91-95.
Mendelssohn H., Marder J.U., Stavу M. Captive breeding of the Houbara and the Development of the Young Birds // Bustard in Decline. Jaipur. 288-292.
Main A. A contribution to the biology of Houbara. 1982-83, wintering population in Baluchistan // Journal of the Bombay Natural History Society. 1984. 81. 537-545.
Mian A., Surahio M.G. Biology of Houbara bustard with reference to Western Baluchistan // Journal of the Bombay Natural History Society. 1983. 80. 111-118.
Platt J. Houbara bustard reseach in Dubai, United Arab Emirates, Bustard Studies. 1985. 3. 101-102.
Surahio M.G. Ecology and distribution of bustards in Sind // Bustard Studies. 1985. 3. 55-58.