




Клейман Д.Г. Социобиология разведения животных в неволе // Биология охраны природы: Пер. с англ. / Под ред. М. Сулея, Б. Уилкокса. / Перевод Остроумова С.А.; Под ред. и с предисл. А.В. Яблокова. – М.: Мир, 1983. С. 275-296.
Для того чтобы разведение животных в неволе шло успешно, необходимо принять во внимание ряд факторов. В их число входят: потребность животных в пространстве, пищевая специализация, требования к параметрам среды (температуре, световому циклу влажности), определенные требования к устройству жилища животных и медицинские вопросы. Необходимые сведения должны быть получены из детального изучения естественных местообитаний. Без этого разведение в неволе будет построено на основе проб и ошибок. Некоторые успешные программы по разведению в неволе содержали в себе элемент беззаботности, вместе с тем многие виды со специфическими, но неустановленными требованиями оказалось невозможным ни разводить, ни содержать в неволе. Исторически сложилось так, что эти виды стали называть «привередливыми». Улучшение методик полевой работы (например, развитие радиотелеметрии) и значительное увеличение размаха и уровня полевых исследований способствовали выявлению особенностей биологии животных. Это привело к тому, что ранее «привередливые» виды стали активно размножаться.
Знание социальных1 требований вида всегда считалось важным для успешного разведения в неволе. Однако поверхностный анализ социальных систем животных может приводить к серьезным ошибкам при организации колоний в неволе. Например, два вида в природе могут жить группами с одинаковым половым и возрастным составом, но иметь совершенно разные системы образования пар и ухода за детенышами, а также разную социальную организацию. Если пытаться к обоим видам в условиях содержания в неволе подходить одинаково, то результатом может быть неудача в содержании и размножении одного из них.
1 Здесь и далее слово «социальный» (social) употребляется в значении, специфическом для описания поведения животных. — Прим. перев.
Более того, иногда виды различаются по способности адаптироваться к условиям, в которых невозможно удовлетворение их специфических требований к пище или жилищу, или по неизвестным еще особенностям социальной лабильности животных, например по степени адаптации животного (без патологических последствий) к непривычной социальной обстановке. Не исключено, что различия между социальными лабильностями разных видов обусловливают, какие именно виды будут процветать в условиях неволи, тем более что информация об их социальных требованиях до сих пор очень скудна.
Социобиология была определена Уилсоном как «систематическое изучение биологической основы социального поведения животных» (Wilson, 1975). Ее корни лежат в современной эволюционной теории, этологии, физиологической психологии, популяционной генетике и экологии. Анализ социального поведения, следовательно, должен включать в себя изучение филогенетической истории вида, экологии, особенностей биологии, популяционной генетики и поведенческих адаптации.
Исследования поведения животных в неволе имеют долгую историю; они содействовали развитию этологии и физиологической психологии. Изучение поведения животных в зоопарках имело свою специфику, связанную с небольшой численностью групп животных и условиями, ограничивающими их передвижение. Поэтому основное внимание уделялось динамике социальных взаимодействий, коммуникации животных, эволюции поведения, анализу мотивации и функций поведения. Некоторые из важных ветвей социобиологии в зоопарках вообще не исследовались. Одна из основных целей этой главы — показать, каким образом социобиология может принести пользу программам по разведению в неволе и как изучение животных в неволе, особенно в зоопарках, может содействовать развитию социобиологии.
СОЦИАЛЬНАЯ ОРГАНИЗАЦИЯ
Социальная организация является собирательным термином, охватывающим ряд характеристик, часть из которых не зависит друг от друга. К социальной организации относятся системы подбора пар для скрещивания у данного вида, будь то моногамия, полигиния, полиандрия или смешанный тип. Стратегии образования пар должны быть дифференцированы в соответствии с тем, сохраняется ли связь данной пары особей, самки и самца, в течение нескольких циклов размножения, или пары каждый раз меняются. Например, многие виды птиц сериально моногамны, у них партнеры меняются каждый сезон; у других моногамных видов пары образуются на длительное время или даже на всю жизнь.
Стратегия отношений с молодняком тоже влияет на конечную социальную организацию вида. Наиболее важны следующие три момента; 1) степень контакта между матерью к ювенильной особью; 2) степень удержания самкой воспитываемых ювенильных особей вместе; 3) степень участия самцаили старших неразмножающихся потомков в родительском уходе.
Численность группы частично подвержена воздействию стратегий образования пар и выращивания молодняка. Но на численность группы влияют также и другие факторы: привычки питания, распределение корма и способ его получения, требования к укрытию, поведение, связанное с защитой от хищников, а также такие филогенетические ограничения, как размер тела и подвижность.
Успешное содержание вида в неволе зависит от знания деталей его социальной организации. Например, средний размер группы, половой и возрастной состав стаи волков (Canislupus) и прайда львов (Pantheraleo) могут быть сходными. Однако система подбора пар, особенности ухода за детенышами и социальные организации этих хищников коренным образом отличаются (Kleiman, Eisenberg, 1973). Эти различия имеют большое значение для разработки программ по разведению в неволе этих двух видов. Волки в основном моногамны. Как правило, размножается только одна пара — основатели группы. Взрослый самец и старший детеныш помогают выращивать младших детенышей, но последние подавлены в репродуктивном отношении. Самки волка так же активны в конкуренции, как и самцы (возможно, даже более), и могут входить в серьезные конфликты после достижения половой зрелости. По-видимому, самки так же, как и самцы, уходят из группы, в которой родились (Kleiman, Brady, 1978).
В группе львов размножаются сразу несколько самцов и несколько самок. Самцы являются обычно братьями и должны конкурировать с другими, неродственными самцами за контроль в прайде. Ядром последнего являются родственные самки и их потомки. Для львов характерна постоянная смена самцов в прайде от года к году. Молодые самцы уходят из родного прайда, а самки обычно остаются и размножаются в рамках той группы, где они родились. Взрослые самцы не участвуют в родительском уходе за детенышами (Schaller, 1972; Bertram, 1976).
В неволе описанные выше отличия в поведении волков и львов могут проявляться на разных стадиях развития размножающейся группы. Стаю волков лучше всего основать с помощью пары неродственных особей. Прайд львов, по-видимому, можно создать сразу из группы родственных, терпимо относящихся друг к другу самцов и группы родственных самок. Взрослеющие самцы и самки в волчьей стае могут войти в конфликт с отцом и матерью соответственно, тогда как среди львов такие конфликты развиваются в основном только между самцами. У волков начало репродуктивного периода обычно сопровождается драками особей одного пола; у львов это наблюдается значительно реже. У львов при выращивании детенышей несколько самок обычно объединяются, тогда как у волков такое объединение менее вероятно. Для решений, принимаемых в связи с разведением животных в неволе, существенное значение имеет каждый из рассмотренных выше факторов.
Стратегия подбора партнеров для размножения
Обычно определения этой стратегии в некоторой степени упрощают реальную ситуацию. Дженни (Jenni, 1974) пришел к выводу, что самцы и самки могут иметь различные стратегии в рамках одной системы размножения в зависимости от организации размножения во времени. Если у самки млекопитающего короткий эструс и внутри социальной группы в данное время в размножении участвует только один самец [например, у оленей Давида (Elaphurusdavidianus) или гамадрилов (Раpiohamadryas)], то самка может рассматриваться как моногамная в ее отношениях с самцом, а самец рассматривается как полигинный. Если основной участвующий в размножении самец меняется ежегодно или каждые два года, то самка может рассматриваться как сериально моногамная. Из анализа подобных временных изменений структуры размножающейся популяции становится ясно, что при любом определении системы скрещивания нельзя исключать фактор времени.
Организация размножения во времени крайне важна для правильного ухода за колонией животных в зоопарке, поскольку она имеет потенциальное генетическое значение. Например, самец оленя Давида в течение сезона размножения конкурирует за положение хозяина гарема. В течение гона, по-видимому, в размножении участвует почти исключительно один самец. Однако другие самцы могут достичь статуса хозяина гарема, когда доминирующий самец утомлен. Главенствующее положение приобретается путем схваток между самцами, что может вести — и ведет — к гибели некоторых содержащихся в неволе особей (Wemmer, 1977; Wemmer, Collins, готовится к печати). Один и тот же самец может удерживать главенствующее положение в гареме в течение ряда лет, но в конце концов ему приходится уступать кому-то из более молодых самцов, достигших своего расцвета. Поэтому для предотвращения конкуренции между самцами и недопущения естественной элиминации пока менее сильных самцов разумной политикой является изоляция в течение гона большинства самцов и допуск к копуляции с самками лишь одного-двух самцов. Потенциальная возможность гибели животных в результате схваток велика. В связи с этим, несмотря на то что целью любой программы по длительному разведению в неволе должно быть созданиетаких условий, при которых все-таки идет отбор самцов, обычно руководители программы и сотрудники, ухаживающие за данной группой животных, сами решают, какие именно особи будут размножаться.
Процесс отбора самцов происходит не только путем схваток; иногда смена самца может иметь крайне разрушительные последствия для всей социальной организации группы. Например, у некоторых видов лангуров (Presbytisssp.) в односамцовых устоявшихся стаях смена самца происходит периодически (Rudran, 1973; Sugiyama, 1965; Blaffer-Hrdy, 1977). Это же явление было описано и для других мартышковых обезьян и лангуров (Struhsaker, 1977). Возможно, оно существует и у других видов приматов, имеющих тенденцию жить в стаях, где самцы ранжируются по возрасту, и в односамцовых стаях (Eisenberg et al., 1972). Смены самцов обычно сопровождаются большим социальным переворотом и напряжением, включая гибель детенышей, частично в результате инфантицида со стороны вторгающегося самца или самцов (Rudran, 1973; Blaffer-Hrdy, 1977).
Рудран (Rudran, 1973) отметил, что у красномордого ланrypa (Presbytissenex) такие смены самцов наблюдаются в среднем каждые три года. Вместе с тем, по оценкам Блаффер-Хрди (Blaffer-Hrdy, 1977), эти смены наблюдались в одной популяции Presbytisentellusраз в 28 месяцев. Одним из результатов таких смен является предотвращение инбридинга внутри дема.
Ясно, что было бы неприемлемо поддерживать такие изменения в колонии лангуров, содержащихся в неволе, т. е. разрешать группе полувзрослых и взрослых самцов вторгаться в установившиеся размножающиеся группы. Более правильным в этих условиях было бы заменять взрослого самца через каждые два-три года другим самцом. Однако даже такие планируемые замены могут приводить к гибели ювенильных особей и детенышей вследствие агрессии со стороны интродуцированного самца; при этом в зоопарках повышалась бы смертность ювенильных особей. Разумным решением данного вопроса может быть временная отмена интродукции нового самца, т. е. воздержание от размножения до окончания того периода, когда большинство детенышей и ювенильных особей в группе кормятся грудью. В любом случае план ухода за видами с описанной выше репродуктивной стратегией должен учитывать, что при замене самцов каждый третий или четвертый год может отсутствовать когорта молодых особей.
Есть сведения о случаях, когда в группах приматов, содержащихся в зоопарках, самцы и другие члены группы убивали детенышей. Точное описание всех обстоятельств, связанных с этим, включая информацию о том, у каких именно видовнаблюдается инфантицид, может помочь в дальнейшем совершенствовании ухода за животными. Такое описание было бы полезно для выяснения роли и эволюции инфантицида в диких популяциях приматов.
Казалось бы, вполне посильно искусственно ограничить конкуренцию между особями одного пола и при этом сохранить путем манипуляции особями или группами связанные с такой конкуренцией преимущества аутбридинга. Однако есть одна дополнительная трудность: некоторые виды или особи не могут успешно размножаться без конкуренции. У самцов некоторых млекопитающих, например у макак-резусов(Масаса mulattа), есть сложная система обратной связи, регулирующей уровень андрогенов посредством социального поведения (Rose et al., 1975). Эта система может воздействовать на силу либидо у самцов. Социально доминантные формы поведения (например, угрозы и схватки) и победы в агрессивных дуэлях играют существенную роль в достижении гормональных уровней, адекватных для успешного сексуального поведения. Отсутствие соперничества может приводить к снижению либидо самцов и в результате к подавлению размножения.
Сходному воздействию могут быть подвержены и самки. Отсутствие соперничества между самцами, например, ведет к уменьшению у них уровня андрогенов, а это в свою очередь снижает репродуктивную функцию усамок. Подобное явление — просто более общий случай эффекта Виттена, который присущ самкам мышей, содержащимся в группе без самца. У таких самок замечена нерегулярность или даже исчезновение репродуктивных циклов. Последние восстанавливаются, если самки мышей видят готового к размножению самца или ощущают его запах (Whitten, Bronson, 1970). Более того, эстральный цикл самок может быть нерегулярным или подавленным в присутствии самца, моча которого или другие выделения содержат мало андрогенов.
Большинство зоопарков знакомо с такими ситуациями, когда имеется единственная пара, которая упорно не размножается, несмотря на все попытки создать для нее подходящие условия обитания, обеспечить ей соответствующее питание и удовлетворить другие потребности. В настоящее время в Национальном зоологическом парке США содержится единственная пара равнинных горилл (Gorillagorilla) и пара индийских носорогов (Rhinocerusunicornis), которые не размножаются, возможно, потому, что у них нет соперничества между самцами. У обоих видов наблюдали единственную успешную попытку к спариванию, за которой последовало подавление эстрального цикла самок и связанное с ним угасание интереса со стороны самца.
Успешное разведение моногамных млекопитающих является проблемой для большинства зоологических институтов. Во-первых, в природе довольно сложно распознать, какая именно стратегия подбора пар имеется у каждого конкретного вида, поскольку особи моногамных видов могут образовывать группы численностью от одного до пятнадцати животных (рис. 1; Kleiman, 1977; 1980). Например, насекомоядные рыжие прыгунчики (Elephantulusrufescens) склонны жить поодиночке (Rathbun, 1979), тогда как размножающаяся группа гиеновых собак (Lycaonpictus) весьма велика по численности: в нее входят полувзрослые и взрослые «помощники» и 6-7, а иногда до 15 детенышей (Kleiman, Eisenberg, 1973). Оба этих вида раньше считались полигамными, пока подробные полевые исследования не вскрыли истинную систему образования пар у них (Kuhme, 1965; van Lawick, 1973; Frame et al., в печати; Rathbun, 1979).
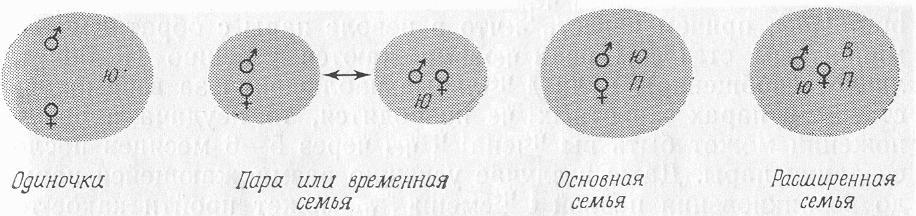
РИС. 1. Размер групп и их структура у некоторых моногамных млекопитающих. Круги представляют общую территорию. Расположение символов размножающихся самцов, размножающихся самок, ювенильных потомков (Ю), полувзрослых потомков (П) и родственных взрослых особей (В) внутри круга указывает на расстояние между особями. В условиях «пары» молодые особи могут жить со взрослыми лишь временно, до своего ухода из группы. В группе может быть более чем один член, отмеченный символом Ю, П или В, поэтому численность группы может достигать 10-15 особей.
При разведении моногамных видов необходимо учитывать тот факт, что в каждой группе размножается обычно не более чем одна самка. Поэтому следует поддерживать сразу несколько пар или семейных групп, чтобы гарантировать размножение даже в случае гибели активного в половом отношении животного, самца или самки. Для этого требуются дополнительные площади в клетках и внимание со стороны сотрудников, ухаживающих за животными; необходимо позаботиться также о соответствующей численности групп и их половом и возрастном составе.
На размножение моногамных животных могут отрицательно влиять особенности, связанные с подбором совместимых пар и соперничеством особей одного пола. Хотя при создании новой пары вполне вероятно, что они будут размножаться, при неблагоприятном стечении обстоятельств животные могут оказаться несовместимыми друг с другом. Например, у рыжих прыгунчиков самки обычно доминируют над самцами (Rathbun, 1979; причем известно, что в неволе пары с обратным доминантным статусом редко размножаются успешно (Rathbun, личное сообщение). Если детальные наблюдения за взаимодействием в парах животных не проводятся, то неудача в размножении может быть выявлена лишь через 5-6 месяцев после создания пары. Даже в случае успешно размножающейся пары до возникновения первой беременности может пройти какое-то время.
У некоторых видов селекцию пар можно стимулировать, если создать такие условия, когда одна особь выбирает другую между двумя потенциальными партнерами. Однако это возможно лишь в случае, когда взрослые особи одного пола содержатся вместе и не вступают в противоборство. У золотистой (или львиной) игрунки (Leontideusrosalia), угрожаемого вида приматов из юго-восточных прибрежных дождевых лесов Бразилии, это легче провести в случае, когда особями одного пола являются самцы (рис. 2). В ходе изучения процесса подбора партнеров были проведены наблюдения над десятью тройками, состоящими из взрослой или только что достигшей зрелости самки и двух самцов. Из этих десяти троек только одну пришлось расформировать из-за слишком сильной агрессивности между самцами (рис. 3, тройка G). Однако в большинстве этих троек сексуальное поведение проявлял только один из самцов (рис. 3). Сексуальное поведение у неактивных в половом отношении самцов в тройках Е и I часто состояло из садок без полного их завершения, и поэтому копуляция была, естественно, безрезультатной. В случае тройки G, которая распалась, самка забеременела от самца I вскоре после окончания наблюдений. В случае тройки J самка забеременела от самца I вскоре после образования тройки, хотя за время наблюдений садок не было замечено.

РИС. 2. Моногамные львиные (золотистые) игрунки (Leontideusrosalia) — угрожаемый вид приматов из юго-восточных дождевых лесов прибрежной части Бразилии. В родительском уходе, помогая переносить малышей, участвуют самцы и другие родственники. (Фото Национального зоологического парка.)
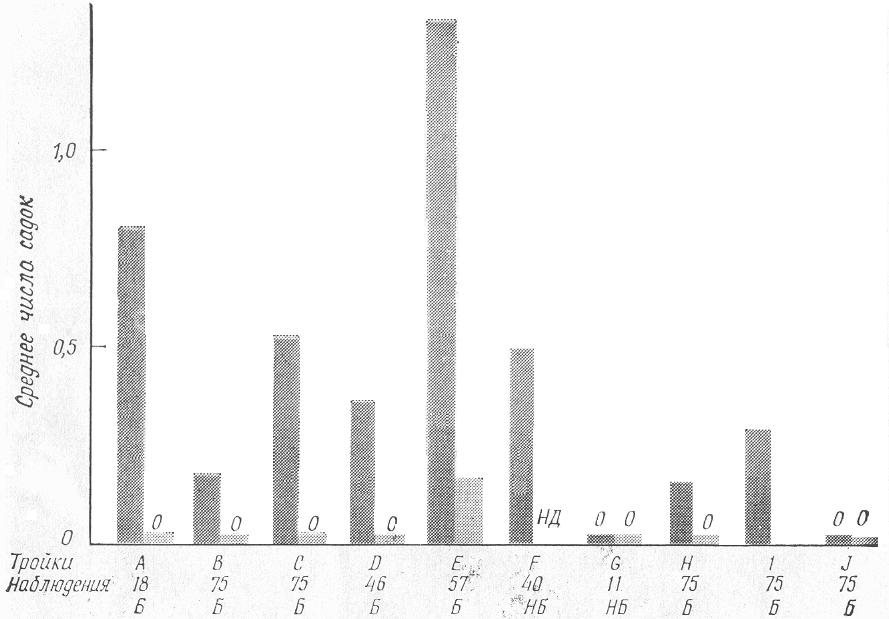
РИС. 3. Частота садок каждого самца в десяти тройках (A—J) львиной игрунки, состоящих из двух самцов и одной самки. НБ — нет беременности, Б — беременность. Цифры внизу обозначают число получасовых сеансов наблюдения за данной тройкой. В отношении самца 2 из тройки Fданных получить не удалось. В каждой паре столбиков левый (темно-серый) относится к самцу 1, а правый (светло-серый) — к самцу 2.
В этом исследовании были получены и другие интересные результаты. Во-первых, явные признаки доминирования у самцов проявлялись лишь изредка; сексуально неактивные самцы не изолировались от пары. Так, все особи одной тройки отдыхали и спали вместе. Во-вторых, имелись лишь минимальные свидетельства того, что самка сознательно выбирала одного из самцов. На рис. 4 приведены подробности груминга (взаимной чистки шерсти) при взаимодействии самки и самцов в десяти тройках. В большинстве случаев сексуально активный самец чистил шерсть самке значительно больше, чем сексуально неактивный. В трех из четырех исключений самка была родственной сексуально неактивному самцу (в тройках Н и F — сестра, в тройке J — мать) и жила вместе с ним и другими членами семьи до того, как была создана тройка. Лишь в четырех тройках самка избирательно чистила шерсть самцу, с которым она спаривалась; в двух из этих четырех троек самкабыла сестрой сексуально неактивного самца. Таким образом, способность самки предпочитать определенного самца сильнее проявляется, когда самка выбирает между родственником и неродственником.
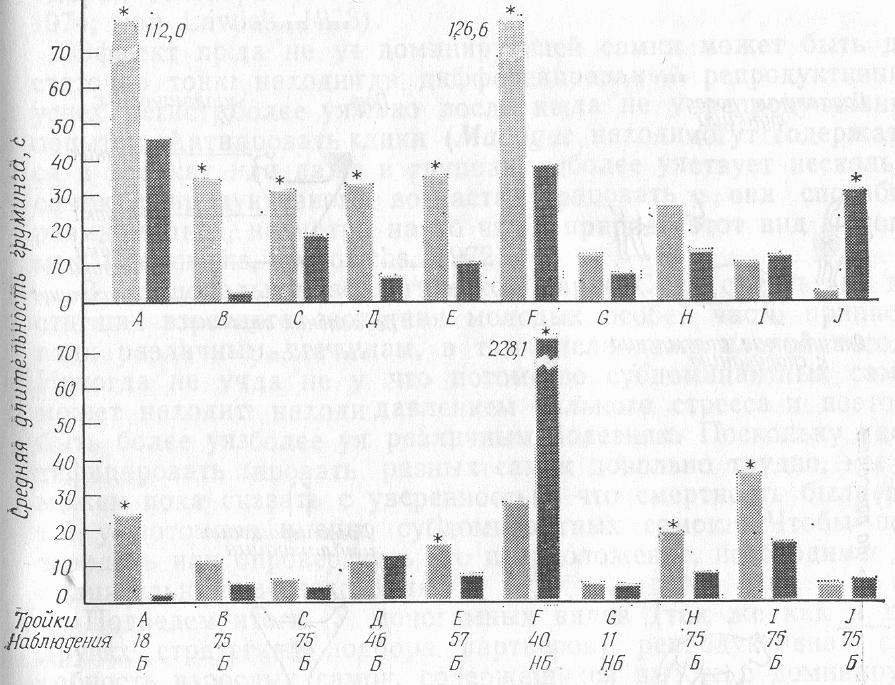
РИС. 4. Груминг в десяти тройках (А—J) львиной игрунки. Верхняя часть рисунка — чистка шерсти самок самцами, нижняя — чистка шерсти самцов самками. В каждой паре столбиков левый (светло-серый) относится к сексуально активному самцу, правый (темно-серый) — к сексуально неактивному. Обозначения внизу, под горизонтальной линией, как на рис. 3. Звездочка обозначает существенную разницу во времени между чисткой шерсти самок самцами и чисткой шерсти самцов самками (тест Вилкоксона для ранжирования спаривающихся пар).
Отсутствие открытого соперничества самцов и ярко выраженного выбора у самок имеет некоторый смысл в плане социальной организации групп золотистых игрунок. Их семейные группы состоят из полувзрослых и, возможно, взрослых особей младшего поколения, которые помогают выращивать детенышей — носят их на себе, участвуют в кормлении. Терпимое отношение к родственникам (пока они не пытаются изменить порядок доминирования при размножении) является преимуществом для размножающейся пары, поскольку помощники способствуют увеличению выживаемости ее потомков. Таким образом, неразмножающиеся полувзрослые и взрослые особи включены в деятельность семьи и не подвергаются агрессивным выпадам со стороны доминирующей размножающейся пары.
У некоторых моногамных млекопитающих соперничество особей одного пола может быть более сильным не среди самцов, а среди самок (Kleiman, 1981). Соперничество может выражаться в обычной манере (например, в драках), но дифференциальный успех при размножении наблюдается и без бросающегося в глаза антагонистического поведения; благодаря этому успеху поддерживается репродуктивное превосходство единственной самки. Различные способы, используемые самкой для поддержания доминирования при размножении, свидетельствуют о том, насколько тонкими являются некоторые из его механизмов (рис. 5).

РИС. 5. Поведенческие и физиологические механизмы, вовлечённые в подавление размножения подчинённых самок у видов млекопитающих с выраженной моногамией. Для разных видов характерен тот или иной механизм подавления. Подавление размножения наблюдается и у млекопитающих с другими системами подбора партнёров.
У обыкновенных игрунок (Callithrixjacchus) нормальный репродуктивный цикл имеет только одна взрослая самка в группе. В семьях или искусственных группах эстральный цикл у всех самок, кроме доминирующей, подавлен (Неаrn, 1977; Lunn, 1978). У волков (Canislupus) доминирующая самка может предотвратить копуляцию самца с субдоминантной самкой путем яростных угроз и атак, которые препятствуют установлению партнерских отношений (Rabb et al., 1967). Если же субдоминантная самка моногамного вида забеременеет, то стрессовое состояние, в котором она находится вследствие совместной жизни с доминирующей самкой, может вызвать аборт или рождение мертвого плода. Эта форма ингибирования при размножении несколько раз была отмечена у листоноса Пратта (Myoproctapratti).
Наконец, даже когда субдоминантная самка успешно спаривается, беременеет и рожает, выживание ее потомков находится под угрозой со стороны доминирующей самки. И для волков, и для гиеиовых собак сообщалось, что доминирующие самки убивали детенышей второй самки или настолько разрушали взаимоотношения матери с детенышем, что последний умирал от заброшенности, голода или беспокойства (Altmann,. 1974; van Lawick, 1973).
Эффект присутствия доминирующей самки может быть достаточно тонким, и тогда дифференциальный репродуктивный успех регистрируют только после нескольких репродуктивных етопыток. Антилопы дикдики (Madoguakirki) могут содержаться в тройках или даже в группах, где присутствует несколько самок репродуктивного возраста, причем все они способны размножаться, несмотря на то что в природе этот вид моногамен (Hendrichs, Hendrichs, 1972).
В Национальном зоологическом парке США смерть не достигших взрослого состояния молодых особей часто приписывали различным причинам, в том числе даже плохой погоде. Никогда не учитывали, что потомство субдоминантных самок может находиться под давлением сильного стресса и поэтому быть более уязвимым к различным болезням. Поскольку идентифицировать потомков разных самок довольно трудно, мы не можем пока сказать с уверенностью, что смертность была выше у потомков именно субдоминантных самок. Чтобы подтвердить или опровергнуть это предположение, необходимы дополнительные исследования.
Подведем итоги. У моногамных видов (так же как и при других стратегиях подбора партнеров) репродуктивная способность взрослых самок, содержащихся вместе с доминирующими самками, может быть подавлена посредством множества механизмов, включая: нарушение или подавление эстрального цикла, предотвращение спаривания, нарушение беременности, гибель детенышей на разных постнатальных стадиях. Репродуктивное подавление может вызываться не только прямым действием доминирующей самки, но с помощью тонких механизмов одним ее присутствием. Эти механизмы могут приводить к стрессу субдоминантной самки, негативно воздействовать на беременность и лактацию, а также вызывать стресс у ее детенышей, тем самым увеличивая их подверженность заболеваниям и уменьшая вероятность их выживания.
По-видимому, в использовании форм репродуктивного подавления у самок, как полигинных, так и моногамных млекопитающих имеются видоспецифические тенденции (Eisenberg, 1967). Изучение этих форм у разных видов должно принести огромную пользу для понимания биологии видов и улучшения содержания млекопитающих в неволе. При исследовании этих вопросов нельзя забывать о том, что репродуктивное подавление может быть опосредовано всего лишь одним обонянием. Например, у некоторых видов обонятельные контакты с репродуктивно доминантными соседями могут быть достаточными для подавления размножения пары, особи которой являются во всех отношениях подходящими друг ддя друга. Предполагали, что это имело место у львиных игрунок и тамаринов, правда, данное предположение не было строго доказано. Обзор данных по ольфакторному контролю репродуктивных процессов недавно сделал Доти (Doty, 1976).
В обсуждение стратегий подбора партнеров необходимо включать и анализ того, в какой мере каждому виду в природе присущ инбридинг. У многих млекопитающих скрещивание между особями старшего и младшего (дочернего) поколения (отец — дочь, мать — сын) предотвращается двумя следующими факторами: 1) уходом молодых особей до того, как они достигнут половой зрелости, и 2) исчезновением, смертью или старением родителей до момента достижения потомком половой зрелости. В зоопарках эти природные факторы часто не действуют из-за специфики содержания в неволе. При этом есть примеры, показывающие, что существует подавление скрещивания между близкородственными особями. Например, в стае волков, ведущих свое происхождение из одного выводка и содержащихся вместе с 1963-1964 гг. в Лондонском зоопарке, не было случаев беременности самок вплоть до 1973 г. Когда же появились детеныши, то в первых двух пометах ни один из них не выжил (Olney, 1975; 1976). У самок ежегодно наблюдалась течка, но успешных копуляций не замечали, несмотря на то что стая была организована обычным образом, с иерархией среди самцов и самок (Kleiman, неопубликованные наблюдения, 1964-1969).
В трех тройках львиных игрунок (Leontideusrosalia), где самка была родственна одному или двум самцам (как сестра или мать), спаривание происходило только с неродственными самцами, несмотря на то что все родственные самцы были половозрелыми (рис. 3; Kleiman, 1978). Майнарди (Mainardi, 1963а, b, с) показал, что молодые самки мышей предпочитают партнерство с незнакомыми особями, а не с теми, с кем они выращивались. Эти факты свидетельствуют о существовании поведенческого механизма, подавляющего инбридинг. Ясно, что степень такого подавления у разных видов варьирует, причем подавление может быть преодолено в отсутствие подходящего партнера. Но эффект подавления может ощущаться в программах по разведению в неволе, если для размножения доступны только родственные особи. В любом случае инбридинга следует по возможности избегать.
Численность и структура группы
Знание стратегии скрещивания данного вида позволяет ответить на вопрос, какое соотношение особей разного пола при содержании в неволе является оптимальным. Однако знание стратегии скрещивания не всегда дает достаточную информациюо пространственных или социальных требованиях данного вида. Например, рассмотрим крошечных мартышек (Cercopithecus). Они полигамны, и тем не менее самцы и самки предпочитают обособляться в однополые подгруппы; размножение идет слабо, если особей разного пола насильно заставлять тесно контактировать друг с другом. Самки становятся крайне агрессивными, а самцы могут умереть от чрезмерного беспокойства (Rowell, 1973). Следовательно, в условиях нехватки площадей в вольерах этих мартышек надо содержать в двух изолированных однополых группах, за исключением брачного периода.
Как уже отмечалось (рис. 1), у моногамных млекопитающих численность групп разных видов может значительно различаться. Поэтому при планировании ухода за ними и планировании их содержания имеет значение, может ли семейнаягруппа долго сохраняться социально стабильной. Например, разведение в неволе рыжих прыгунчиков наиболее успешно, когда отнятых от груди потомков изолируют от родителей еще до момента рождения следующего поколения. Напротив, это не является необходимым и, даже более того, нежелательно вслучае волков или африканских диких собак. В некоторых ситуациях необходимость изоляции размножающегося самца, самки или пары особей не очевидна, за исключением того случая, когда нет успешного размножения. И гепард (Acinonyxjubatus), и красная панда (Ailurusfulgens) могут содержаться в неволе как группы особей, не проявляющих агрессивного поведения. Однако у гепардов размножение наиболее успешно, когда самка постоянно находится в изолированном состоянии, за исключением брачного периода, а самки красной панды размножаются лучше всего в условиях, когда они либо изолированы, либо содержатся в одном помещении с единственным самцом.
Существует некоторая информация о наборе видов, особи которых для успешного размножения нуждаются в изоляции — поодиночке или парами. Однако очень мало известно о том, какие виды в целях стимулирования размножения следует содержать в группах — хотя бы с минимальной численностью. Например, многие летучие мыши обычно размножаются в больших колониях. В двух малых группах летучей мыши длинноязыкового листоноса (Glossophagasoricina), включавших только по одной самке, размножение шло вяло. У родственного ему вида летучей мыши, короткохвостого листоноса (Carolliа perspicillata), две колонии, в которых было 10-20 самок, размножались в тех же условиях среды успешно. Однако в малой группе Carollia, где было только 3-4 самки, размножение было нестабильным (Kleiman, не опубликовано).
Необходимость присутствия в группе минимального числасамок обусловлена несколькими причинами. У некоторых видов летучих мышей колониальное гнездование служит механизмом сохранения тепла. Это особенно важный фактор для обитающих в умеренной зоне обыкновенных летучих мышей, хотя он имеет значение и для тропических летучих мышей. Известно, что у нетопыря-карлика (Pipistrelluspipistrellus) температура воздействует на длительность беременности (Raсеу, 1973), причем лактация и рост молодых особей тоже могут зависеть от температуры. Живя в колонии, самка и ее детеныши могут поддерживать оптимальную температуру с небольшими энергетическими затратами.
Тот факт, что самки некоторых видов размножаются в больших группах, может быть связан со стратегией защиты ют хищников. Часто это сопровождается синхронизацией репродуктивной активности. Например, синхронная рождаемость отмечена у антилопы гну (Connochaetestaurinus), живущей в Восточной Африке. Было показано, что это адаптация, благодаря которой создается избыток (по отношению к хищникам) потенциальных жертв (детенышей антилопы гну), в результате чего снижается общая смертность молодняка (Estes, 1976). Хотя нет данных, которые указывали, бы на то, что размножение антилопы гну в неволе снижается при числе самок менее какого-то минимального, тем не менее могут существовать такие виды, у которых эволюция синхронизации репродуктивной активности как стратегии защиты от хищников продвинулась так далеко, что для адекватного размножения в группе необходима большая численность самок.
Стратегии выращивания молодняка
Если стратегии выращивания молодняка не уделяется достаточного внимания, то выживание потомков может уменьшаться. Самки многих млекопитающих выращивают детенышей совместно. Пока нельзя сказать, в какой степени изоляция лактирующей самки влияет на развитие детеныша. Например, у многих видов обезьян-тонкотелов ювенильные особи вскоре после рождения передаются «теткам» (Blaffer-Hrdy, 1976; Ноrwich, Manski, I975) и после этого проводят с матерями лишь ограниченное время. Неизвестно, какое из двух следующих возможных объяснений правильно — то ли матери просто терпимы к участию других животных в выращивании их детенышей, то ли эта стратегия выращивания настолько специализирована,, что детеныш, содержащийся в неволе совместно с одной изолированной самкой, будет иметь мало шансов на выживание. У нас пока просто нет данных, чтобы судить о том, имеется ли в неволе повышенная смертность детенышей у таких животных, как слоны, дельфины и обезьяны-тонкотелы, yкоторых «тетки» являются составной частью системы выращивания молодняка.
Сходная проблема существует и у видов, у которых выращивать детенышей помогают взрослые самцы и молодые помощники. Янтшке (Jantschke, 1973) получил доказательства того, что для успешного выращивания щенков самками кустарниковой собаки (Speothosvenaticus) необходимо присутствие отца. Не вызывает сомнения, что сходные явления будут найдены и у других моногамных экзотических видов. Дадли изящно показал. (Dudley, 1974), что самцы мышевидного хомячка (Peromyscuscalifornicusparasiticus) вносят вклад в выживание детенышей, согревая их в отсутствие самки. Самцы могут оказывать и другую помощь: они кормят детенышей, защищают от хищников и переносят их (Kleiman, 1977).
У львиной игрунки взрослые самцы носят молодняк на себе и делятся с ним пищей, а также приучают его к общественному образу жизни (Hoage, 1977; 1978; рис. 2). Львиные игрунки обычно имеют одного - трех детенышей. Когда задались целью определить с какого дня после рождения детенышей отцы начинают их носить (табл. 1), пришли к выводу, что отцы начинают раньше проявлять родительскую заботу, если приплод более многочисленный. Таким образом, степень родительской опеки со стороны отцов и других родственников и время ее проявления, по-видимому, зависят от нужд матери в данный момент времени.
Таблица 14
Среднее число дней после рождения,
спустя которое самец львиной игрунки начинает носить детенышей, в случае различной численности помета
(данные по четырем самкам)
|
Численность помёта |
|||
|
1 |
2 |
3 |
|
|
Среднее |
14,2 |
8,6 |
3 |
|
Диапазон |
12-17 |
5-11 |
- |
|
Число выводков |
6 |
9 |
1 |
У многих млекопитающих самки выращивают своих детенышей в изоляции от других особей своего вида. Насильственный контакт или содержание в тесноте, как было показано, уменьшают у некоторых видов выживаемость молодняка. Мартин (Martin, 1968, 1975) первым обнаружил, что самка тупайи (Tupaiabelangeri), содержавшаяся вместе с единственным самцом, нуждалась в отдельном гнездовом ящике для детеныша.
У этих животных система родительского ухода относится к типу «абсенти» (от англ. "absentee"1; Eisenberg, 1977): если самка вынуждена жить с детенышем и самцом в одном гнезде, детеныш подвергается каннибализму. Сходным образом размножение у красных панд (Ailurusfulgens) почти всегда безуспешно, если две или большее число самок содержатся вместе. Хотя самки становятся беременными, детеныши неизбежно игнорируются или их убивают. Стратегия подбора пар у красных панд неизвестна, но данные, подобные упомянутым, наталкивают на предположение, что красные панды в природе моногамны.
1«Absentee» (англ.) - лицо, длительное время проживающее вне своего постоянного местожительства. — Прим. перев.
ПРОБЛЕМА РАССЕЛЕНИЯ
Один из наименее разработанных вопросов в области оптимального поддержания численности популяций в неволе - как и когда отселять из группы взрослое или молодое животное, сохраняя при этом оптимальную половую и возрастную структуру группы и не разрушая ее социальной динамики. Особенности биологии групп в природе (их образование, поддержание и распад) изучены слабо, а между тем все они могут оказаться очень важными в понимании популяционного регулирования данного вида (Eisenberg et al., 1972).
В полевых исследованиях многих млекопитающих основное внимание было сконцентрировано на долговременных изменениях групп известного состава, и информация об особенностях биологии групп только начинает появляться. Однако уже показано, что формы расселения и распада групп отличаются у разных видов и едва ли могут быть предсказаны на основе знаний о социальной организации или стратегии образования пар. Более того, Бекофф (Bekoff, 1977) высказал предположение, что поведение особей одного вида в отношении стратегии расселения различается. Он ссылается на наблюдения индивидуальных отличий в выводках койотов (Canislatrans), где самые доминантные и самые подчиненные из животных в выводке в наименьшей мере взаимодействуют с сибсами. Бекофф предполагает, что невзаимодействующие особи наиболее склонны уходить нз родной группы, хотя причины их отделения от остальной части выводка совершенно различны.
Некоторые наблюдения за львиными игрунками свидетельствуют о том, что предсказывать, какие животные, вероятнее всего, покинут группу (и, следовательно, должны быть удалены из семейной группы), довольно трудно. Молодые самцы львиных игрунок еше в семейной группе начинают проявлять такие признаки половой зрелости, как запаховое мечение (Kleiman, Mack, в печати) и демонстрационное поведение (Rathbun, в печати). Наоборот, самки не проявляют признаков половой зрелости до тех пор, пока их не удалили от родителей и не поселили в паре с неродственным самцом; таким образом, в семейной группе они социально и репродуктивно ингибированы. Однако отношения доминирования существуют у обоих полов и у сибсов одного пола могут быть идентифицированы. Отношения доминирования у самцов-сибсов иногда включают в себя драки, приводящие к царапинам и небольшим ранам. Молодой доминирующий самец может преследовать самку старшего поколения, а также активнее, чем отец, осуществлять запаховое мечение и демонстративное поведение; при этом родители его терпят, и наблюдаются лишь небольшие ссоры.
Что касается доминирующих самок дочернего поколения, отмечены 2 случая их гибели, причем в обеих семейных группах они были убиты при прямом участии их матерей. Небезынтересно, что атакованные самки даже не успели достичь половой зрелости (обе имели возраст около года) и обе имели сестру-близнеца, либо находились в семейной группе, где было еще несколько других самок дочернего поколения. Среди молодых доминирующих самцов гибель никогда не регистрировалась. Случайный наблюдатель семейных групп мог бы рекомендовать устранение созревающих самцов из-за их повышенной половой активности в группе. Однако на самом деле самки подвергаются большему риску уже на ранних стадиях развития. Вероятно, таких самок нужно отсаживать из семейной группы, чтобы они не были убиты. У львиных игрунок соперничество между самцами носит не очень опасный для их жизни характер и разрешается преимущественно посредством ссор без серьезного ущерба для здоровья.
ВОЗДЕЙСТВИЕ СОДЕРЖАНИЯ В НЕВОЛЕ НА ПОВЕДЕНИЕ
При изучении животных в неволе или при попытках экстраполировать данные об их поведении на поведение диких животных (или в обратном направлении) надо учитывать долговременные и краткосрочные воздействия содержания в неволе на поведение. Начиная с самого первого выбора — каких именно животных взять для содержания в зоопарках,— человек ведет селекцию на содержание и разведение особей с определенными поведенческими фенотипами (например, неагрессивных и покорных). Особи, которые проявляют либо крайний страх, либо крайнюю агрессивность по отношению к человеку, обычно исключаются из размножения, поскольку с ними слишком трудно справиться. Такой отбор со стороны человека может быть бессознательным.
Независимо от отбора человеком особи, легко поддающиеся стрессу, не способны успешно размножаться; поэтому они почти не вносят вклад в генетический фонд. Не исключено, что в неволе происходят изменения в поведенческом генотипе популяции, например уменьшается тенденция избегать людей и хищников.
В условиях зоопарков могут сохраняться такие генотипы, которые вряд ли бы выжили в природе. При заботе со стороны человека вырастают ослабленные детеныши, которые вносят в зоопарковые популяции такие гены, которые в природе должны элиминироваться. Классический пример этого — тигры (Pantherа tigris), частичные альбиносы, полученные путем инбридинга в нескольких зоопарках. Неизвестно, имеет ли этот редкий мутант специфические для него поведенческие характеристики; однако у высокоинбредных особей отмечены отклонения от нормы в зрительной системе (Guiilery, Kaas, 1971).
На поведении животных в неволе сказывается то, что молодняк выращивается в необычных условиях среды, в том числе социальной. Выращивание детенышей «на руках» часто впоследствии мешает животным формировать адекватные социальные привязанности. Голдфут (Goldfoot, 1977) пришел к выводу: чем больше степень социальных нарушений у макак-резусов (Масаса mulatta), выращенных на руках, тем ниже конечный репродуктивный успех. Есть сообщения и о других сходных корреляциях между нарушениями сложных форм поведения взрослых животных и нарушениями в ходе развития. Хищники с помощью родителей и других родственников постепенно учатся обездвиживать и убивать свою жертву. Этот опыт уже трудно приобрести после того, как животное стало взрослым; более того, «неопытная» взрослая особь не в состоянии должным образом обучать своих детенышей. Успех таких методик, как искусственное осеменение и выращивание на руках (при повышенной заботе со стороны человека), способствует увеличению численности популяций видов, содержащихся в неволе, но это происходит за счет изменения нормальных поведенческих реакций. С увеличением зависимости от искусственных средств размножения возможно наступление такой ситуации, когда мы будем сохранять в зоопарках вид, особи которого не способны должным образом ни скрещиваться, ни выращивать молодняк.
Последний фактор, который воздействует на поведение вида в неволе, — это вмешательство человека в нормальный ход как подбора партнеров для скрещивания, так и отделения взрослых или молодых особей. Произвольные решения, основанные на соображениях удобства ухода за животными или другого рода, могут существенно повлиять на генофонд и привести к изменению поведенческих генотипов.
Имеeтся множество факторов, которые оказывают долговременное и краткосрочное воздействие на поведенческий фенотип особей, причем некоторые из них необратимы. Поведенческие изменения могут быть не только приемлемыми, но и желательными для популяций видов, которым суждено сохраняться в неволе неопределенно долго. Однако в зоопарках разводят некоторые виды с намерением вернуть их в конце концов в природу. В этих случаях бессознательное нанесение ущерба генофонду и поведению популяций в неволе может сделать невозможной успешную реинтродукцию. Попытки реинтродукции видов были немногочисленными и большей частью имели ограниченный успех. Существенные детали, которые надо учитывать при таких попытках, обсуждаются Брэмбеллом (Brambell, 1977).
РЕЗЮМЕ
Недостаточно продуманное использование систем скрещивания и выращивания молодняка, а также приемов отделения от размножающейся группы взрослых и молодых особей может отрицательно сказаться на размножении в условиях неволи (Eisenberg, 1967). К сожалению, крайне трудно точно назвать причины неудач в размножении вида — особенно если биология вида изучена недостаточно. В зоопарках редко публикуются результаты неудачных программ по разведению, как это делают, например, патологи, публикуя результаты аутопсии. Исследователи, работающие в зоопарках, не любят обсуждать свои неудачи. Отсутствие публикаций часто связано также с небольшим размером выборки и устным характером данных. Как правило, трудно заключить, в какой мере результаты, полученные на одной особи или группе животных в неволе, характеризуют вид в целом. Например, в зоопарках есть многочисленные случаи инфантицида у приматов, которые прямо сопоставляют со сходными событиями в дикой природе. Однако крайне трудно указать те детали в наблюдениях, сделанных в зоопарках, которые могли бы углубить наше понимание этого феномена, - такие, как половая и возрастная структура группы, а также социальные изменения, которые происходят перед случаями инфантицида. Более того, чтобы определить, у каких видов приматов выше вероятность инфантицида, требуются сравнительные данные, касающиеся предшествующих социальных условий, когда инфантицида не было. Такой анализ возможен, но он требует больших затрат средств и сил.
Интересно, что неудачная программа по разведению в неволе, провалившись, может дать больше для социобиологической теории и знаний, чем успешная программа. Например, поведенческие механизмы, благодаря которым уменьшается вероятность инбридинга, легче анализировать именно в зоопарках. Изучение особенностей биологии тех, видов, у которых подавлено скрещивание между близкими родственниками, может дать ценную информацию, необходимую для нашего понимания предотвращения инбридинга.
Современное знание стратегий скрещивания и выращивания молодняка моногамных игрунок и тамаринов основано на многочисленных неудачах разведения этих приматов в неволе. Зоопарки и другие организации пытались содержать их в искусственно сформированных группах, состоящих из неродственных особей, где было найдено, что размножается не более чем одна самка. Кроме того, было отмечено, что молодые особи, которых отсаживали от родителей после прекращения кормления грудью (но до рождения следующего поколения детей), когда наступала их очередь размножаться, не способны правильно ухаживать за своими детьми. По-видимому, если молодые особи не проходят школу ухода за младшими детенышами, то становясь взрослыми, они не способны адекватно ухаживать за собственными детьми (Hoage, 1977; 1978).
Исследования видов в неволе вносят свой вклад в социо-биологию различным образом. Длительные наблюдения за животными в зоопарках дают важную информацию о социальной динамике групп в течение нескольких поколений. Такой подход, был реализован при изучении волка (Rabb et al., 1967), а недавно - оленя Давида (Wemmer, Collins, готовится к печати).
К сожалению, подобные исследования в зоопарках проводятся редко, поскольку они длятся десятилетиями или более — особенно в случае долгоживущих видов. Однако некоторые из наиболее важных проблем в социобиологии относятся к взаимодействию родственных особей и изучению того, в какой степени генетическое родство особей влияет на их общественное поведение, а такие проблемы могут эффективно изучаться в популяциях животных, содержащихся в зоопарке.
РЕКОМЕНДУЕМАЯ ЛИТЕРАТУРА
CrandallL.S. 1964. The Management of Wild Mammals in Captivity, University of Chicago Press. Chicago.
Eisenberg J.F. 1966. The Social Organization of Mammals. In: Handbuch der Zoologie, VIII (10/7), Lieferung 39, De Gruyter, Berlin.
Hediger H. 1950. Wild Animals in Captivity, Butterworth's and Co., Ltd., London.
Hediger H. 1955. Psychology and Behaviour of Captive Animals in Zoos and Circuses, Butterworth's and Co., Ltd., London.
Hediger H. 1969. Man and Animal in the Zoo, Seumour Lawrence/Delacorte Press, New York.
Research in Zoos and Aquariums, 1975, Institute of Laboratory Animal Resources, National Academy of Sciences, Washington, D.С
International Zoo Yearbook, Volumes 1-17, Zoological Society of London,. 1960-1977.
Wilson E.O. 1975. Sociobiology: The New Synthesis, Harvard University Press, Cambridge.
Der Zoologische Garten, Leipzig, East Germany.
ЛИТЕРАТУРА
Altmann D., 1974. Beziehungen zwischen sozialer rangordnung und jungenaufzucht bei Canis lupus L., Zool. Gart. N. F. Jena, 44, 235-23
Bekoff M., 1977. Mammalian dispersal and the ontogeny of individual behavioral phenotypes, Amer. Nat., III, 715-732.
Bertram B., 1976. Kin selection in lions and evolution. In: Growing Points in Ethology, P.P.G. Bateson, R.A. Hinde (eds.), Cambridge University Press. Cambridge, pp. 281-301.
Blafjer-Hrdy S., 1976. The care and exploitation of nonhuman primate infants by conspecifics other than the mother. In: Advances in the Study of Behavior, Vol. 6, J. S. Rosenblatt et al. (eds.), Academic Press, New York, pp. 101-158.
Blaffer-Hrdy S., 1977. The Langurs of Abu, Harvard University Press, Cambridje, Massachusetts.
Brambell M.R., 1977. Reintroduction, Inter. Zoo. Yearb., 17, 112-116.
Doty R.L., 1976. Mammalian Olfaction, Reproductive Processes and Behavior, Academic Press, New York.
Dudley D., 1974. Contributions of paternal care to the growth and development of the young in Peromyscuc californicus, Behav. Biol., 11, 155-166.
Eisenberg J.F., 1967. A comparative study in rodent ethology with emphasis on evolution of social behavior, I, Proc. U.S. Nat. Mus., 122, 1- 51.
Eisenberg J.F., 1977. The evolution of the reproductive unit in the class -Mammalia. In: Reproductive Behavior and Evolution, J.S. Rosenblatt and B.R. Komisarik (eds.), Plenum Publishing Co., New York, pp. 39-71.
Eisenberg J.F., Muckenhirn N.A., Rudran R.A., 1972. The relation between ecology and social structure in primates, Science, 176, 863-874.
Estes R.D., 1976. The significance of breeding synchrony in the wildebeest, E. Afr. Wildl. J., 14, 135-152.
Goldfoot D.A., 1977. Rearing conditions which support or inhibit later sexual potential of laboratory monkeys: hypothesis and diagnostic behaviors, Lab-Anim. Sci., 27, 548-556.
Guillery R.W., Kass J.H., 1971. Genetic abnormality of the visual pathway in a "white" tiger, Science, 180, 1287-1289.
Hearn J.P., 1977. The endocrinology of reproduction in the common marmoset, Callithrix jacchus. In: The Biology and Conservation of the Callitrichidae, D.G. Kleman (ed.), Smithsonian Institution Press, Washington D.C., pp. 163-171.
Hendrichs H., Hendrichs U., 1972. Dikdik und Elefanten: Okologie und Soziologie zweier Afrikanischer Huftiere, R. Piper and Co., Munich.
Horwich R., Manski D., 1975. Maternal care and infant transfer in two species of Colobus monkeys, Primates, 16, 49-74.
Hoage R.J., 1977. Parental care in Leontopithecus rosalia: sex and age differences in carrying behavior and the role of prior experience. In: The Biology and Conservation of the Callitrichidae, D.G. Kleiman (ed-), Smithsonian Institution Press, Washington, D.C., pp. 293-305.
Hoage R.J., 1978. Biosocial Development in the Golden Lion Tamarin, Leontopithecus rosalia rosalia (Primates: Callitrichidae), Ph. D. thesis, University of Pittsburgh.
Jantschke F., 1973.On the breeding and rearing of buch dogs, Speothos venaticus, at Frankfurt Zoo, Inter. Zoo Yearb., 13, 141-143.
Jenni D.A., 1974. The evolution of polyandry in birds, Am. Zool., 14, 129-144.
Kleiman D.G., 1977. Monogamy in mammals, Quart. Rev. Biol., 52, 39-69.
Kleiman D.G., 1980. Correlations among life history characteristics of mammalian species exhibiting two extreme forms of monogamy. In: Natural Selection and Social Behavior, R.D. Alexander and D.W.Tinkle (eds.), Chiron Press, New York, 332-344.
Kleiman D.G., Brady C.A., 1978. Coyte behavior in the context of recent canid research: problems and perspectives. In: Coyotes: Behavior and Management, M. Bekoff (ed.), Academic Press, New York, pp. 163-188.
Kleiman D.G., Eisenberg J.P., 1973. Comparisons of canid and felid social systems from an evolutionary perspective, Anim. Behav., 21, 637-659.
Kleiman D.G., Mack D.S., 1980. The effects of age, sex and reproductive status on scent marking frequencies in the golden lion tamarin, Leontopithecus rosalia, Folia Primat, 33, 1-14.
Kuhme W., 1965. Freilandsttidien zur Soziologie des Hyanenhundes (Lycaon pictus lupinsu Thomas, 1902), Z. Tierpsychol., 22, 495-541.
Lunn S.P., 1978. Urinary oestrogen excretion in the common marmoset, Callithrix jacchus. In: The Biology and Behavior of Marmosets, H.J. Wolters and J.P. Hearn (eds.), Eigenverlag Roth, Gottingen. pp. 67-73.
Mainardi D., 1963a. Un esperimento sulla parte attiva svolta dalla femmina nella selexione sessuale in Mus musculus, Arch. Sci. Bio., 47, 227-237.
Mainardi D., 1963b. Speciazone nel topo, fattori etologici determinianti barriere reproduttive tra Mus musculus domesticus e M. m. bactrianus, Instituto Lombardo, Rend. Sc., B., 97, 135-142.
Mainardi D., 1963c. Eliminazione della barriere etologica all’ isolamento riproduttivo tra Mus musculus domesticus e M. m. bactrianus mediante sull’ apprendimento infantile, Instituto Lombardo, Rend. Sc., B, 97, 291-299.
Martin R.D., 1968. Reproduction and ontogeny in the tree-shrews (Tupaia belangeri) with reference to their general behavior and taxonomic relationships, Z. Tierpsychol., 2,5, 409-495, 505-532.
Martin R.D., 1975. Breeding tree-shrews, Tupaia belangeri, and mouse lemurs, Microcebus murinus, in captivity, Inter. Zoo Yearb., 15, 35-41.
Rabb G.B., Woolpy J.H., Ginsburg B.E., 1967. Social relationships in a group of captive wolves, Am. Zool., 7, 305-312.
Racey P.A., 1973. Environmental factors affecting the length of gestation in heterothermic bats, J. Reprod. Fert. Suppl., 19. 175-189.
Rathbun C.D., 1979. Descriptions and analysis of the arch display in the golden lion tamarin, Leontopithecus rosalia rosalia, Folia Primat, 32, 125 -148.
Rose R. M., Bernstein I. S., Gordon T. P., 1975. Consequences of social conflict on plasma testosterone levels in rhesus monkeys, Psychosom. Med., 37, 50-61.
Rowell T.E., 1973. Social organization of wild talapoin monkeys, Am. J. Phys. Anthrop., 38, 593-598.
Rudran R., 1973. Adult male replacement in one male troops of purplefaced langurs (Presbytis senex senex) and its effect on population structure, Folia primat., 19, 166-192.
Schaller G., 1972. The Serengeti Lion, University of Chicago Press, Chicago.
Struhsaker T.T., 1977. Infanticide and social organization in the red tail monkey (Cercopithecus ascanius schmidti) in the Kibala Forest, Uganda, Z. Tierpsychol., 45, 75-84.
Sugiyama Y., 1965. On the social change of Hanuman langurs (Presbyiis entellus) in their natural conditions, Primates, 6, 381-429.
van Lawick H., 1973. Solo, Bantam Books, New York.
Frame L.H., Mahalm I.R., Frame G.W., van Lawick H., 1979. Social organization of African wild dogs (Lycaon pictus) on the Serengeti plains, Tierpsychol., 50, 225-249.
Wemmer C., 1977. Can wildlife be saved in zoos? New Scientist, 75, 585-587.
Wemmer C., Collins L.R., in preparation. Social behavior, social structure and their implications for management. In: The Biology and Captive Management of Pere David's Deer, B. Beck and C. Wemmer (eds.).
Whitten W.K., Bronson F.H., 1970. The role of pheromones in mammalian reproduction. In: Advances in Chemoreception, Vol. 1, J.W. Johnston, D.G. Moulton and A. Turk (eds.), Appleton, New York, pp. 309-325.
Wilson E.E., 1975. Sociobiology; The New Synthesis, Harvard University Press, Cambridge, Massachusets.